

SMRT Regulates Metabolic Homeostasis and Adipose Tissue Macrophage Phenotypes in Tandem
- PMID: 32770234
- PMCID: PMC7478322
- DOI: 10.1210/endocr/bqaa132
SMRT Regulates Metabolic Homeostasis and Adipose Tissue Macrophage Phenotypes in Tandem
Abstract
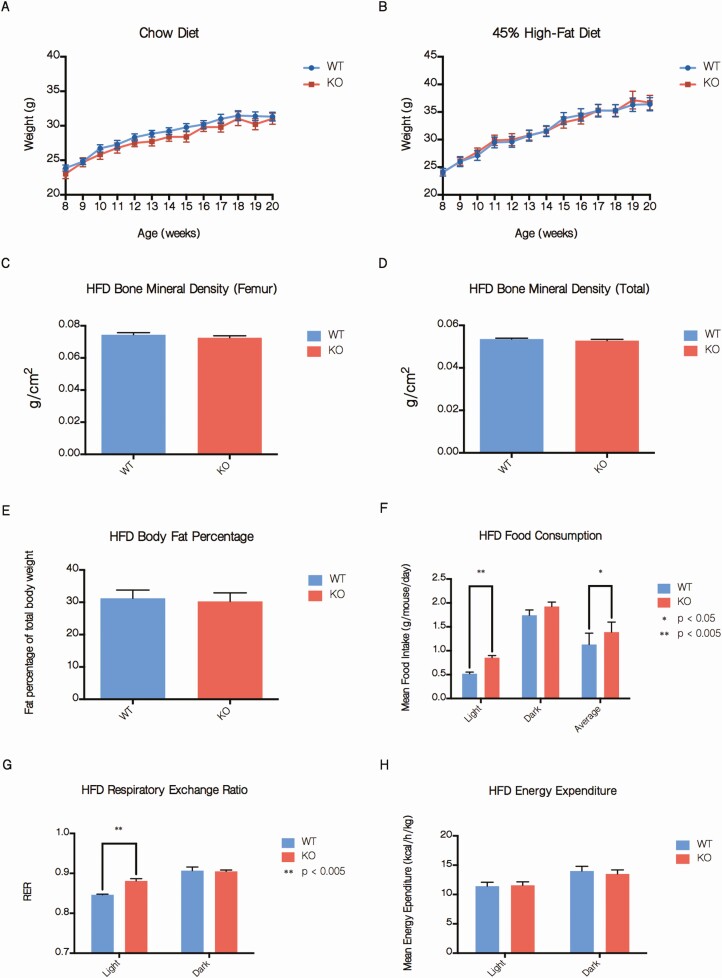
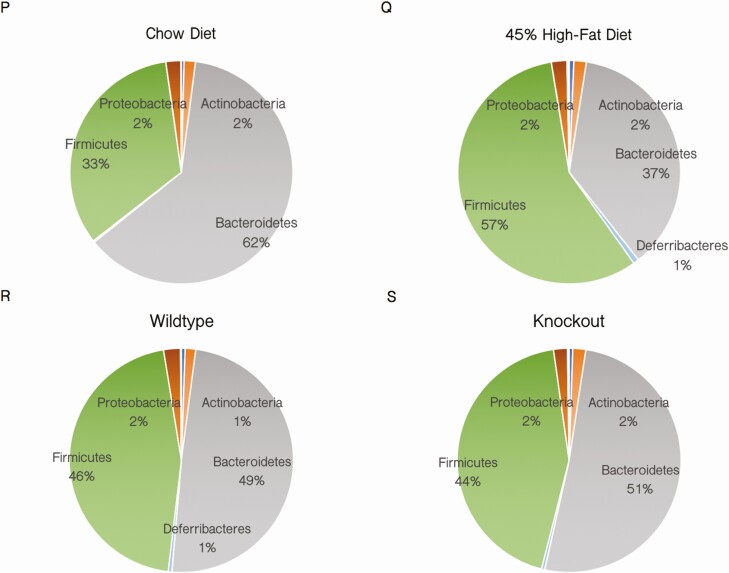
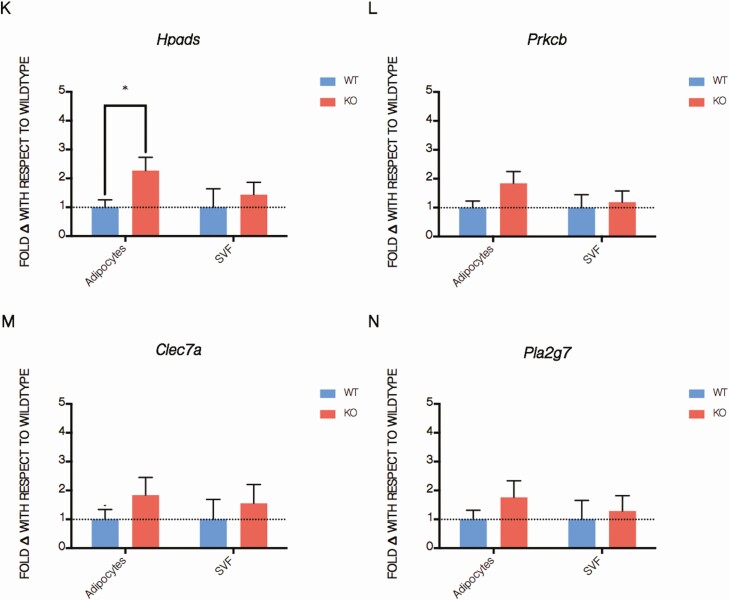
The Silencing Mediator of Retinoid and Thyroid Hormone Receptors (SMRT) is a nuclear corepressor, regulating the transcriptional activity of many transcription factors critical for metabolic processes. While the importance of the role of SMRT in the adipocyte has been well-established, our comprehensive understanding of its in vivo function in the context of homeostatic maintenance is limited due to contradictory phenotypes yielded by prior generalized knockout mouse models. Multiple such models agree that SMRT deficiency leads to increased adiposity, although the effects of SMRT loss on glucose tolerance and insulin sensitivity have been variable. We therefore generated an adipocyte-specific SMRT knockout (adSMRT-/-) mouse to more clearly define the metabolic contributions of SMRT. In doing so, we found that SMRT deletion in the adipocyte does not cause obesity-even when mice are challenged with a high-fat diet. This suggests that adiposity phenotypes of previously described models were due to effects of SMRT loss beyond the adipocyte. However, an adipocyte-specific SMRT deficiency still led to dramatic effects on systemic glucose tolerance and adipocyte insulin sensitivity, impairing both. This metabolically deleterious outcome was coupled with a surprising immune phenotype, wherein most genes differentially expressed in the adipose tissue of adSMRT-/- mice were upregulated in pro-inflammatory pathways. Flow cytometry and conditioned media experiments demonstrated that secreted factors from knockout adipose tissue strongly informed resident macrophages to develop a pro-inflammatory, MMe (metabolically activated) phenotype. Together, these studies suggest a novel role for SMRT as an integrator of metabolic and inflammatory signals to maintain physiological homeostasis.
Keywords: NCoR2; SMRT; adipose tissue; gene knockout; homeostasis; hormone receptor; inflammation; metabolism; nuclear corepressor; transcription regulation.
© Endocrine Society 2020. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures







References
-
- Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab. 2004;89(6):2548-2556. - PubMed
-
- Rezaee F, Dashty M. Role of adipose tissue in metabolic system disorders: adipose tissue is the initiator of metabolic diseases. J Diabet Metab. 2013;13(8):2155–6156.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
