

Influence of the polar light cycle on seasonal dynamics of an Antarctic lake microbial community
- PMID: 32772914
- PMCID: PMC7416419
- DOI: 10.1186/s40168-020-00889-8
Influence of the polar light cycle on seasonal dynamics of an Antarctic lake microbial community
Abstract
Background: Cold environments dominate the Earth's biosphere and microbial activity drives ecosystem processes thereby contributing greatly to global biogeochemical cycles. Polar environments differ to all other cold environments by experiencing 24-h sunlight in summer and no sunlight in winter. The Vestfold Hills in East Antarctica contains hundreds of lakes that have evolved from a marine origin only 3000-7000 years ago. Ace Lake is a meromictic (stratified) lake from this region that has been intensively studied since the 1970s. Here, a total of 120 metagenomes representing a seasonal cycle and four summers spanning a 10-year period were analyzed to determine the effects of the polar light cycle on microbial-driven nutrient cycles.
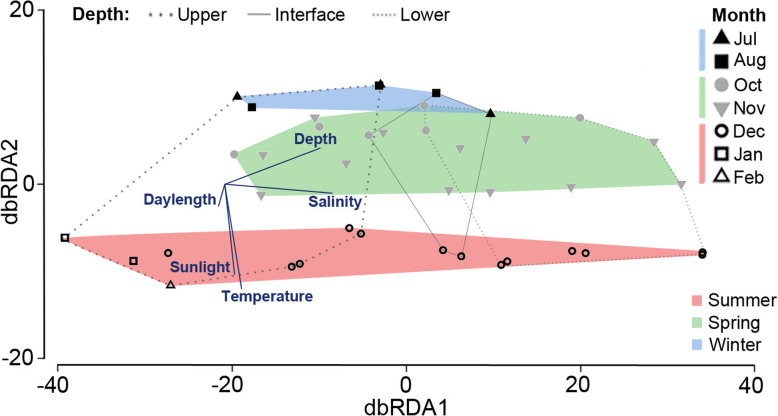
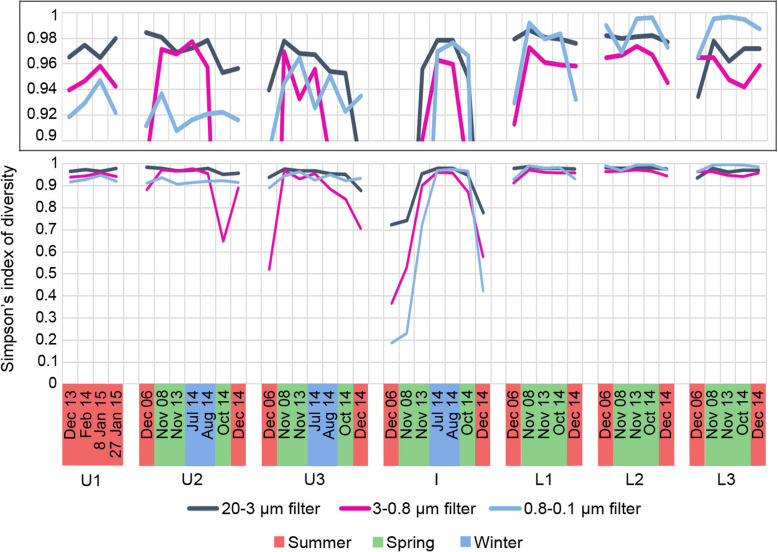
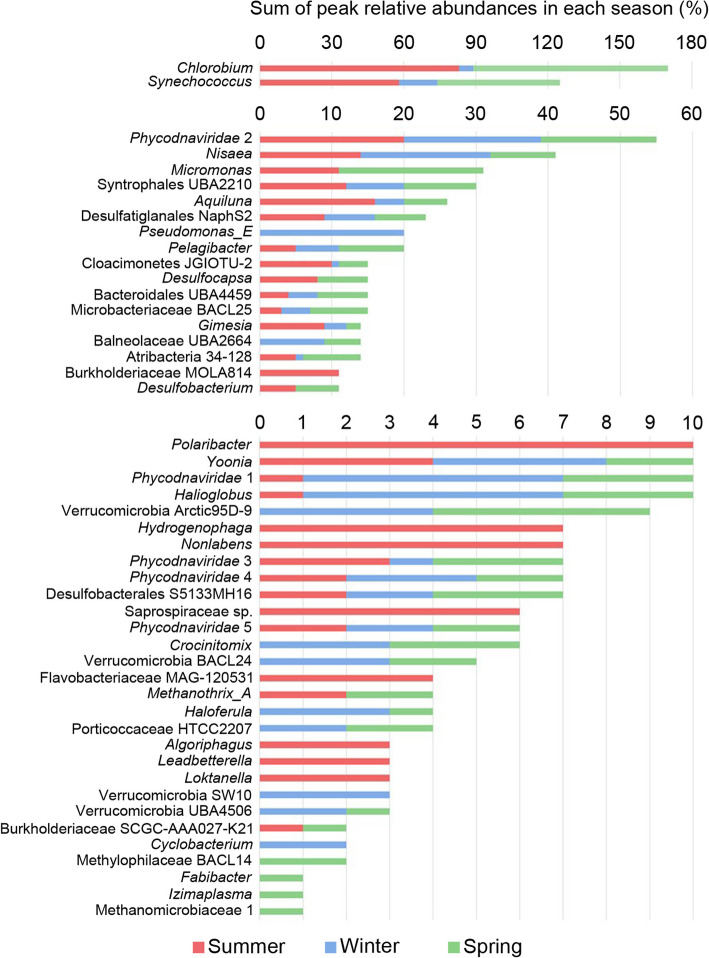
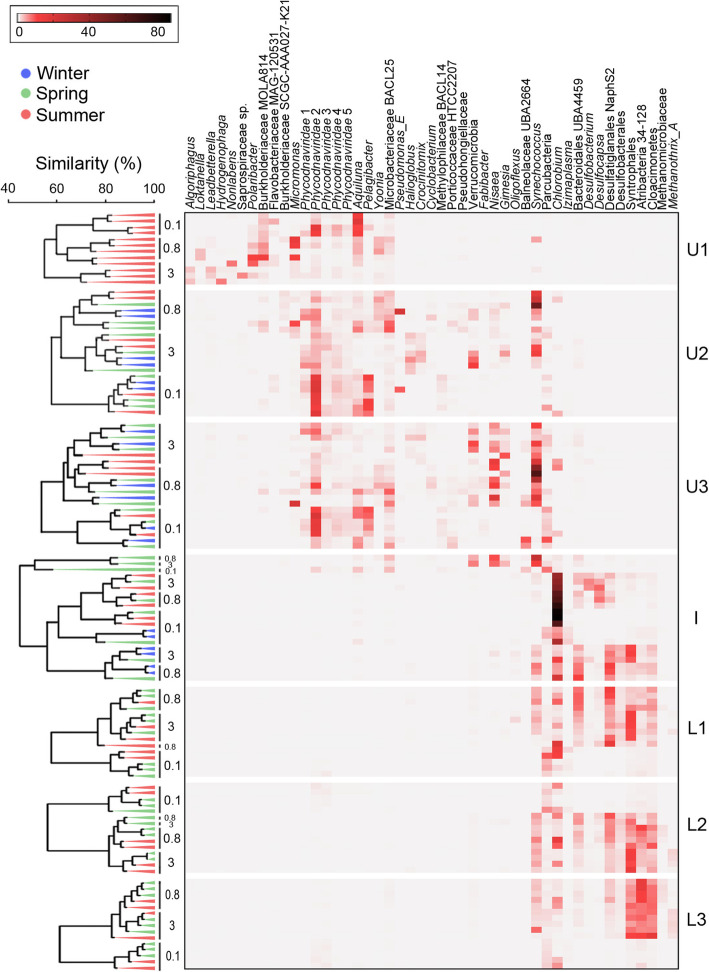
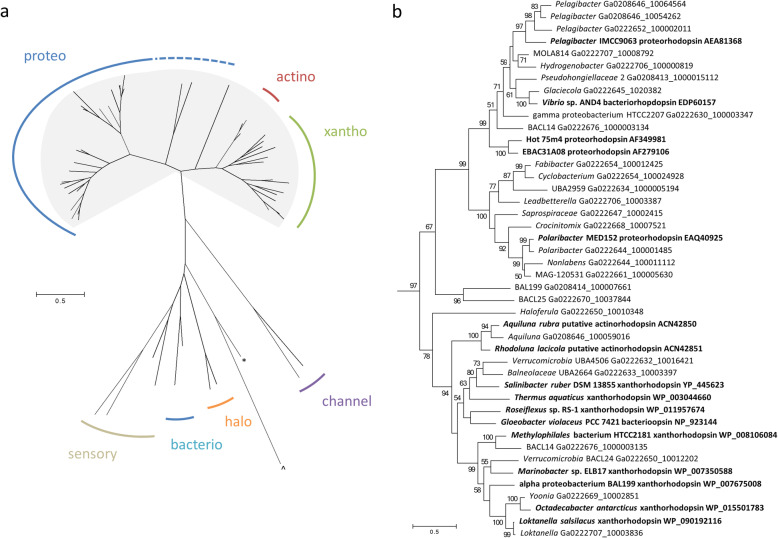
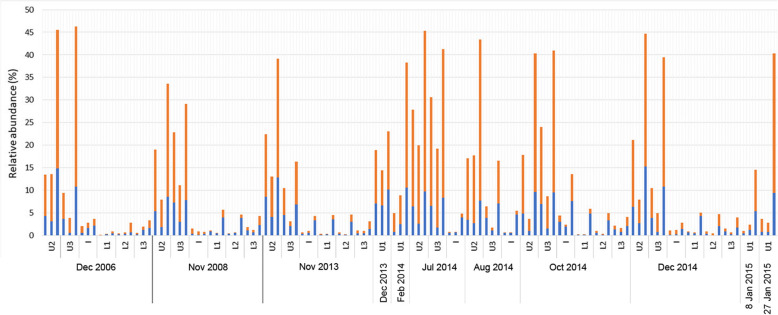
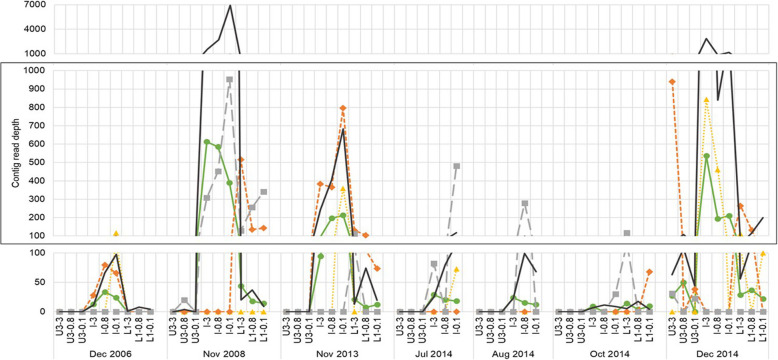
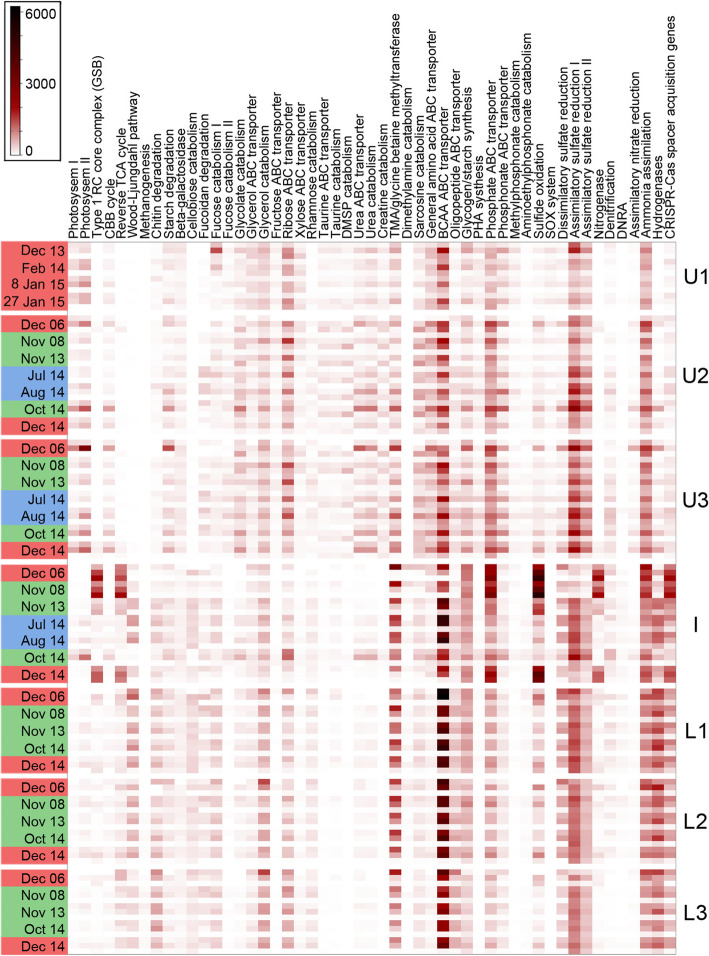
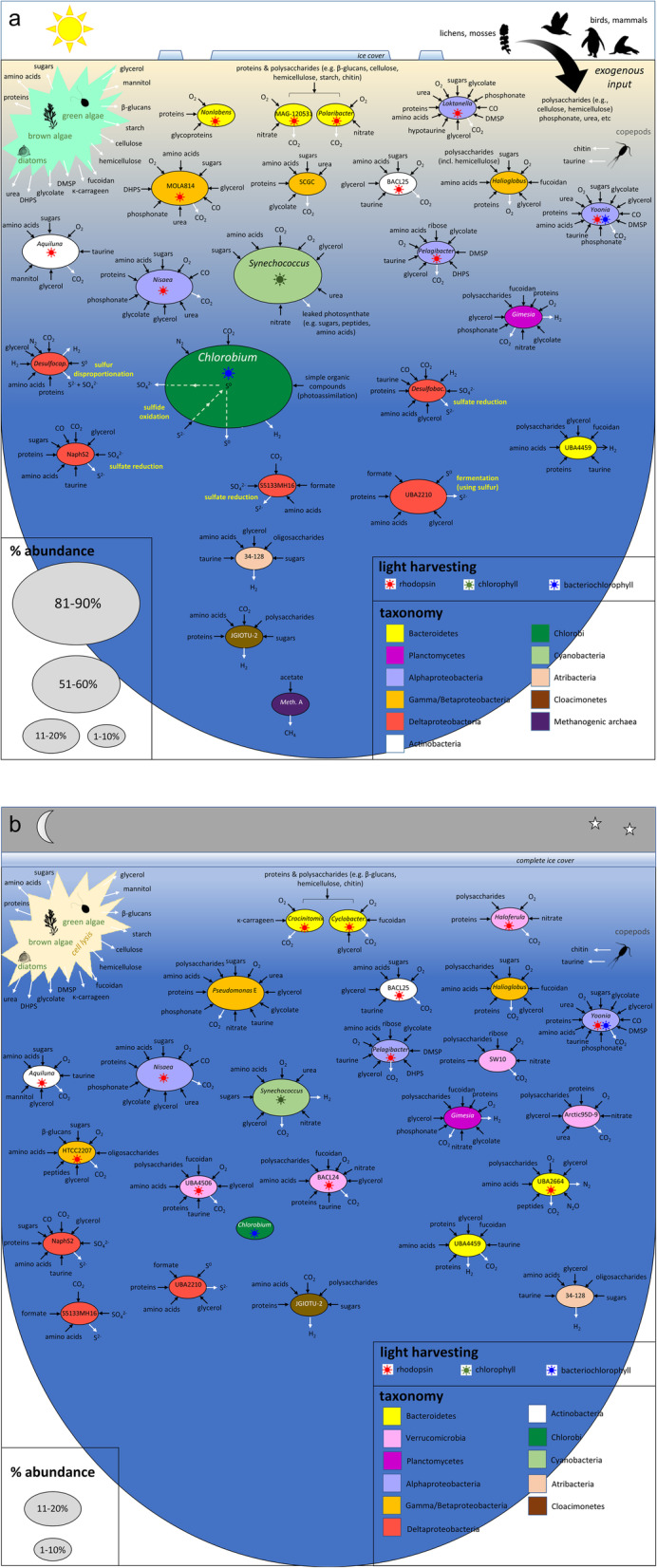
Results: The lake system is characterized by complex sulfur and hydrogen cycling, especially in the anoxic layers, with multiple mechanisms for the breakdown of biopolymers present throughout the water column. The two most abundant taxa are phototrophs (green sulfur bacteria and cyanobacteria) that are highly influenced by the seasonal availability of sunlight. The extent of the Chlorobium biomass thriving at the interface in summer was captured in underwater video footage. The Chlorobium abundance dropped from up to 83% in summer to 6% in winter and 1% in spring, before rebounding to high levels. Predicted Chlorobium viruses and cyanophage were also abundant, but their levels did not negatively correlate with their hosts.
Conclusion: Over-wintering expeditions in Antarctica are logistically challenging, meaning insight into winter processes has been inferred from limited data. Here, we found that in contrast to chemolithoautotrophic carbon fixation potential of Southern Ocean Thaumarchaeota, this marine-derived lake evolved a reliance on photosynthesis. While viruses associated with phototrophs also have high seasonal abundance, the negative impact of viral infection on host growth appeared to be limited. The microbial community as a whole appears to have developed a capacity to generate biomass and remineralize nutrients, sufficient to sustain itself between two rounds of sunlight-driven summer-activity. In addition, this unique metagenome dataset provides considerable opportunity for future interrogation of eukaryotes and their viruses, abundant uncharacterized taxa (i.e. dark matter), and for testing hypotheses about endemic species in polar aquatic ecosystems. Video Abstract.
Keywords: Antarctic microbiology; Green sulfur bacteria; Host-virus interactions; Meromictic lake; Metagenome time series; Microbial food web; Phototroph; Polar light cycle.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Margesin R, Miteva V. Diversity and ecology of psychrophilic microorganisms. Res Microbiol. 2011;162:346–361. - PubMed
-
- Cavicchioli R. Microbial ecology of Antarctic aquatic systems. Nat Rev Microbiol. 2015;13:691–706. - PubMed
-
- Cary SC, McDonald IR, Barrett JE, Cowan DA. On the rocks: the microbiology of Antarctic Dry Valley soils. Nat Rev Microbiol. 2010;8:129–138. - PubMed
-
- Rankin LM, Gibson JAE, Franzrnann PD, Burton HR. The chemical stratification and microbial communities of Ace Lake, Antarctica: a review of the characteristics of a marine-derived meromictic lake. Polarforschung. 1999;66:33–52.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
