

G12/13 is activated by acute tethered agonist exposure in the adhesion GPCR ADGRL3
- PMID: 32778842
- PMCID: PMC7990041
- DOI: 10.1038/s41589-020-0617-7
G12/13 is activated by acute tethered agonist exposure in the adhesion GPCR ADGRL3
Erratum in
-
Publisher Correction: G12/13 is activated by acute tethered agonist exposure in the adhesion GPCR ADGRL3.Nat Chem Biol. 2020 Dec;16(12):1440. doi: 10.1038/s41589-020-0649-z. Nat Chem Biol. 2020. PMID: 32807971
Abstract
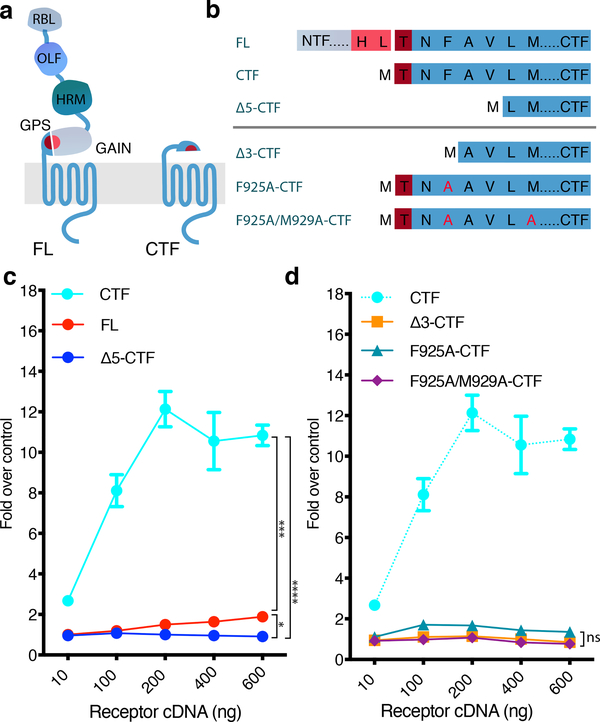
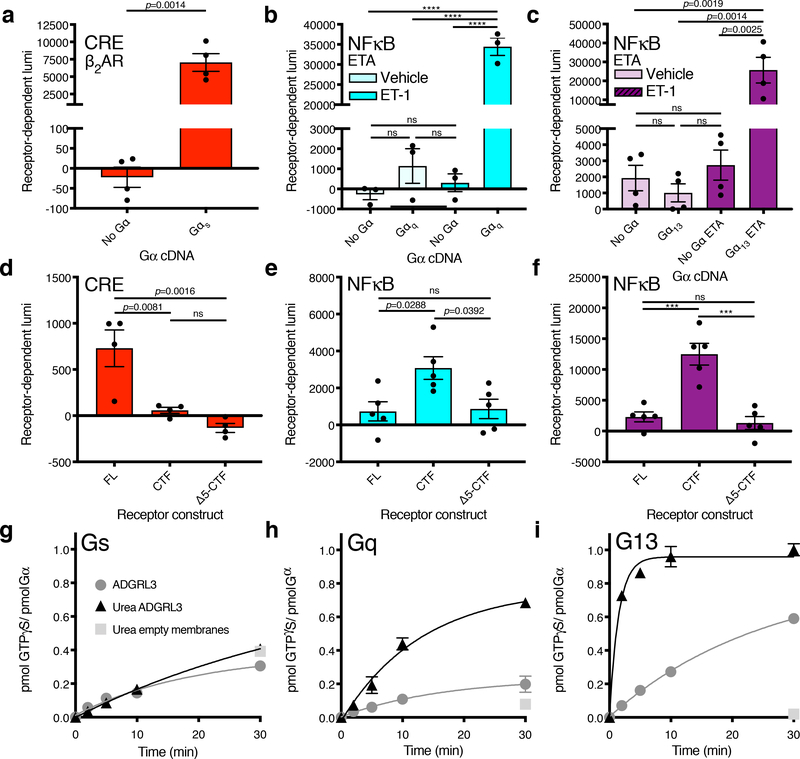
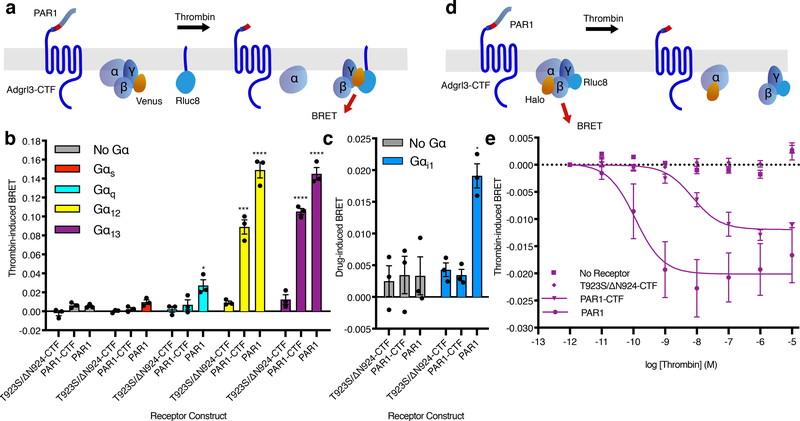
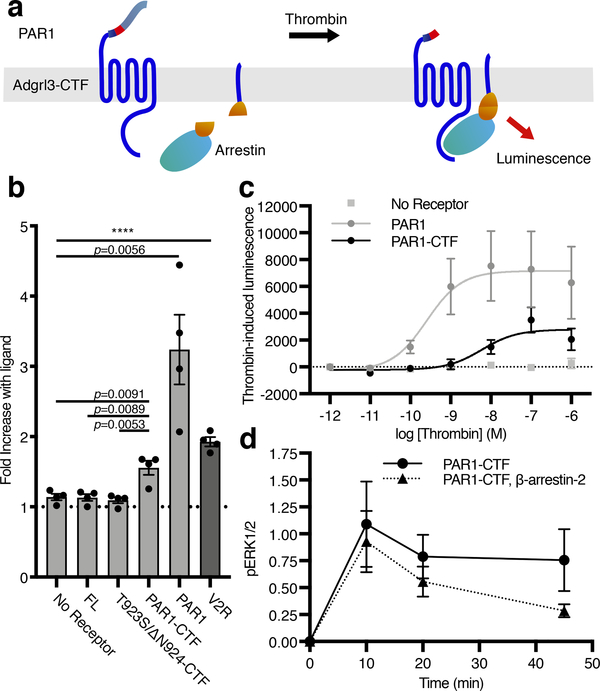
The adhesion G-protein-coupled receptor (GPCR) latrophilin 3 (ADGRL3) has been associated with increased risk of attention deficit hyperactivity disorder (ADHD) and substance use in human genetic studies. Knockdown in multiple species leads to hyperlocomotion and altered dopamine signaling. Thus, ADGRL3 is a potential target for treatment of neuropsychiatric disorders that involve dopamine dysfunction, but its basic signaling properties are poorly understood. Identification of adhesion GPCR signaling partners has been limited by a lack of tools to acutely activate these receptors in living cells. Here, we design a novel acute activation strategy to characterize ADGRL3 signaling by engineering a receptor construct in which we could trigger acute activation enzymatically. Using this assay, we found that ADGRL3 signals through G12/G13 and Gq, with G12/13 the most robustly activated. Gα12/13 is a new player in ADGRL3 biology, opening up unexplored roles for ADGRL3 in the brain. Our methodological advancements should be broadly useful in adhesion GPCR research.
Conflict of interest statement
Competing Interests statement
The authors declare no competing financial interests.
Figures
Comment in
-
Designer adhesion GPCR tells its signaling story.Nat Chem Biol. 2020 Dec;16(12):1280-1281. doi: 10.1038/s41589-020-00673-7. Nat Chem Biol. 2020. PMID: 32989302 No abstract available.
References
References main text
-
- Hamann J, Aust G, Arac D, Engel FB,Formstone C,Fredriksson R,Hall RA,Harty BL, Kirchhoff C, Knapp B, Krishnan A, Liebscher I, Lin HH, Martinelli DC, Monk KR, Peeters MC, Piao X, Promel S, Schoneberg T, Schwartz TW, Singer K, Stacey M, Ushkaryov YA, Vallon, Wolfrum U, Wright MW, Xu L, Langenhan T & Schioth HB. International Union of Basic and Clinical Pharmacology. XCIV. Adhesion G protein-coupled receptors. Pharmacol Rev 67, 338–367 (2015). - PMC - PubMed
-
- Arcos-Burgos M, Velez JI, Martinez AF, Ribases M, Ramos-Quiroga JA, Sanchez-Mora C, Richarte V, Roncero C, Cormand B, Fernandez-Castillo N, Casas M, Lopera F, Pineda DA, Palacio JD, Acosta-Lopez JE, Cervantes-Henriquez ML, Sanchez-Rojas MG, Puentes-Rozo PJ, Molina BSG, Group MTAC, Boden MT, Wallis D, Lidbury B, Newman S, Easteal S, Swanson J, Patel H, Volkow N, Acosta MT, Castellanos FX, de Leon J, Mastronardi CA & Muenke M ADGRL3 (LPHN3) variants predict substance use disorder. Transl Psychiatry 9, 42 (2019). - PMC - PubMed
References online methods
-
- Grundmann M, Merten N, Malfacini D, Inoue A, Preis P, Simon K, Ruttiger N, Ziegler N, Benkel T, Schmitt NK, Ishida S, Muller I, Reher R, Kawakami K, Inoue A, Rick U, Kuhl T, Imhof D, Aoki J, Konig GM, Hoffmann C, Gomeza J, Wess J & Kostenis E Lack of beta-arrestin signaling in the absence of active G proteins. Nat Commun 9, 341 (2018). - PMC - PubMed
-
- Stallaert W, van der Westhuizen ET, Schonegge AM, Plouffe B, Hogue M, Lukashova V,Inoue A, Ishida S, Aoki J, Le Gouill C & Bouvier M Purinergic Receptor Transactivation by the beta2-Adrenergic Receptor Increases Intracellular Ca(2+) in Nonexcitable Cells. Mol Pharmacol 91, 533–544 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
