

Metabolic Heterogeneity and Cross-Feeding in Bacterial Multicellular Systems
- PMID: 32781027
- PMCID: PMC7419710
- DOI: 10.1016/j.tim.2020.03.008
Metabolic Heterogeneity and Cross-Feeding in Bacterial Multicellular Systems
Abstract
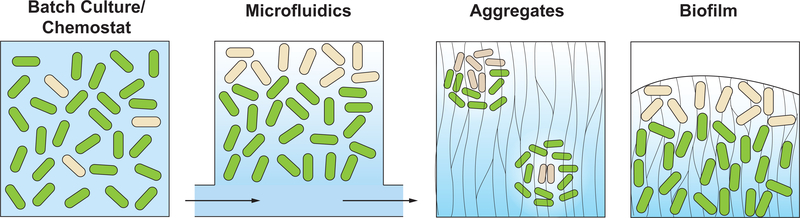
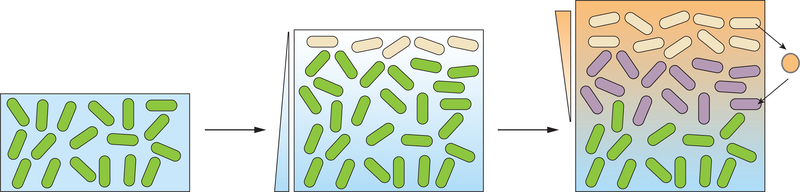
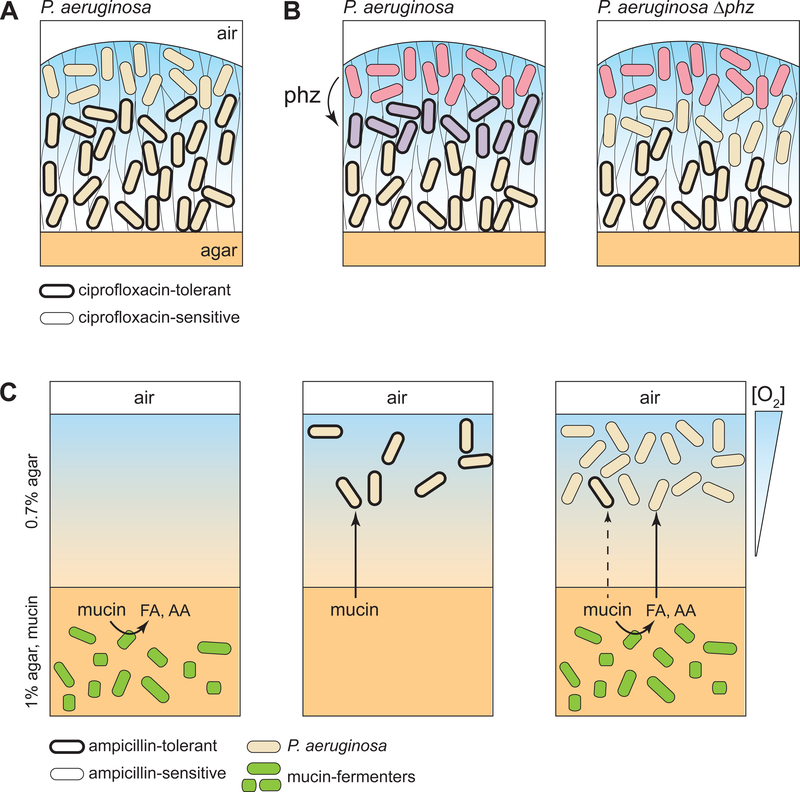
Cells in assemblages differentiate and perform distinct roles. Though many pathways of differentiation are understood at the molecular level in multicellular eukaryotes, the elucidation of similar processes in bacterial assemblages is recent and ongoing. Here, we discuss examples of bacterial differentiation, focusing on cases in which distinct metabolisms coexist and those that exhibit cross-feeding, with one subpopulation producing substrates that are metabolized by a second subpopulation. We describe several studies of single-species systems, then segue to studies of multispecies metabolic heterogeneity and cross-feeding in the clinical setting. Many of the studies described exemplify the application of new techniques and modeling approaches that provide insights into metabolic interactions relevant for bacterial growth outside the laboratory.
Keywords: biofilms; metabolite exchange; microfluidics; phenotypic diversity.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures

References
-
- Batsivari A. et al. (2020) Dynamic responses of the haematopoietic stem cell niche to diverse stresses. Nat. Cell Biol 22, 7–17 - PubMed
-
- Woods HA (2014) Mosaic physiology from developmental noise: within-organism physiological diversity as an alternative to phenotypic plasticity and phenotypic flexibility. J. Exp. Biol 217, 35–45 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
