

Molecular determinants of SR-B1-dependent Plasmodium sporozoite entry into hepatocytes
- PMID: 32782257
- PMCID: PMC7419504
- DOI: 10.1038/s41598-020-70468-2
Molecular determinants of SR-B1-dependent Plasmodium sporozoite entry into hepatocytes
Abstract
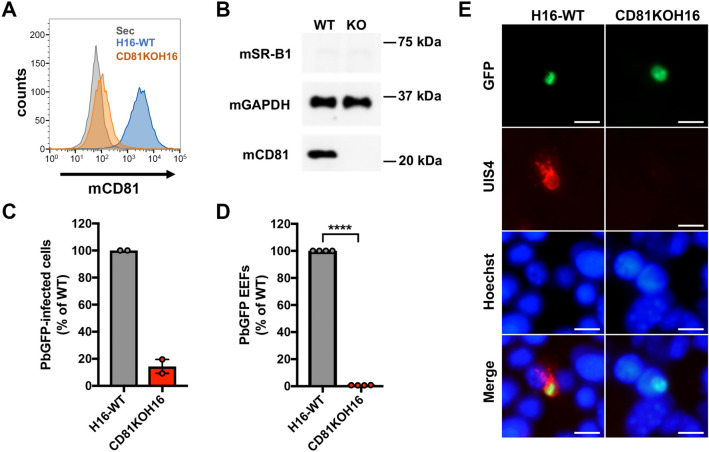
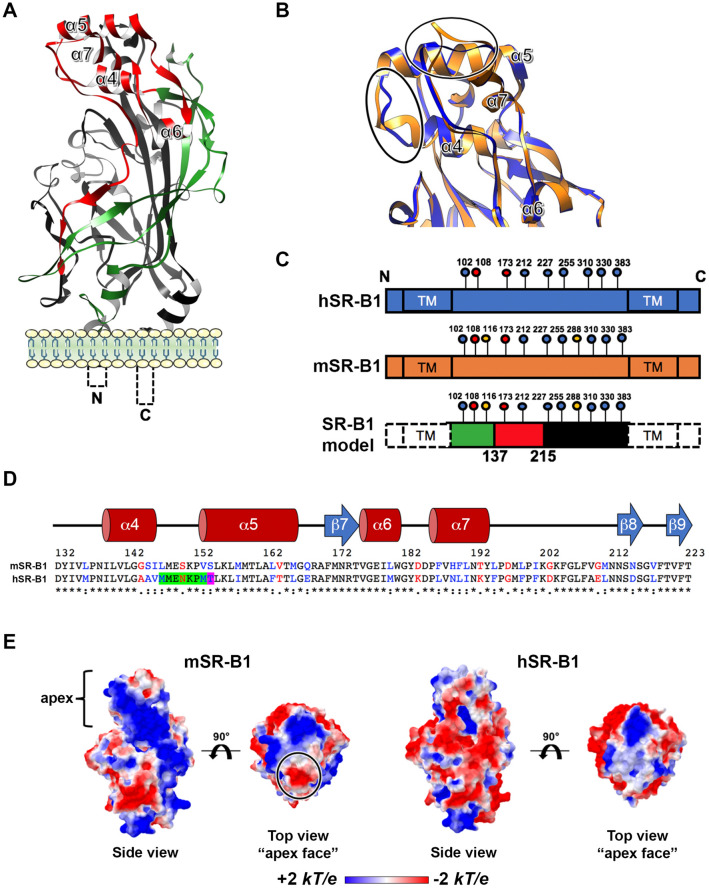
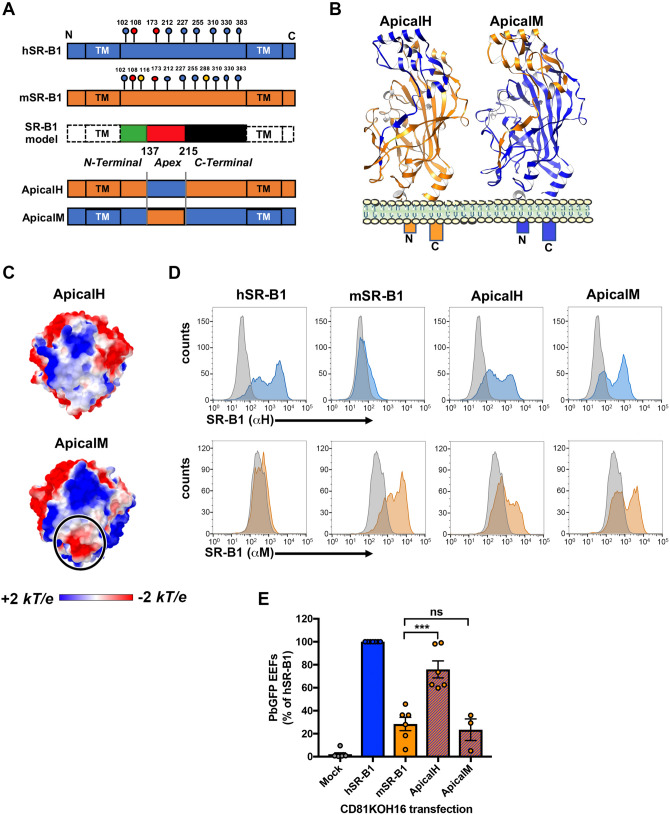
Sporozoite forms of the Plasmodium parasite, the causative agent of malaria, are transmitted by mosquitoes and first infect the liver for an initial round of replication before parasite proliferation in the blood. The molecular mechanisms involved during sporozoite invasion of hepatocytes remain poorly understood. Two receptors of the Hepatitis C virus (HCV), the tetraspanin CD81 and the scavenger receptor class B type 1 (SR-B1), play an important role during the entry of Plasmodium sporozoites into hepatocytes. In contrast to HCV entry, which requires both CD81 and SR-B1 together with additional host factors, CD81 and SR-B1 operate independently during malaria liver infection. Sporozoites from human-infecting P. falciparum and P. vivax rely respectively on CD81 or SR-B1. Rodent-infecting P. berghei can use SR-B1 to infect host cells as an alternative pathway to CD81, providing a tractable model to investigate the role of SR-B1 during Plasmodium liver infection. Here we show that mouse SR-B1 is less functional as compared to human SR-B1 during P. berghei infection. We took advantage of this functional difference to investigate the structural determinants of SR-B1 required for infection. Using a structure-guided strategy and chimeric mouse/human SR-B1 constructs, we could map the functional region of human SR-B1 within apical loops, suggesting that this region of the protein may play a crucial role for interaction of sporozoite ligands with host cells and thus the very first step of Plasmodium infection.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- World Health Organization. World Malaria Report 2019. 1–232 (2019).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
