

Atomistic structure and dynamics of the human MHC-I peptide-loading complex
- PMID: 32788370
- PMCID: PMC7456110
- DOI: 10.1073/pnas.2004445117
Atomistic structure and dynamics of the human MHC-I peptide-loading complex
Abstract
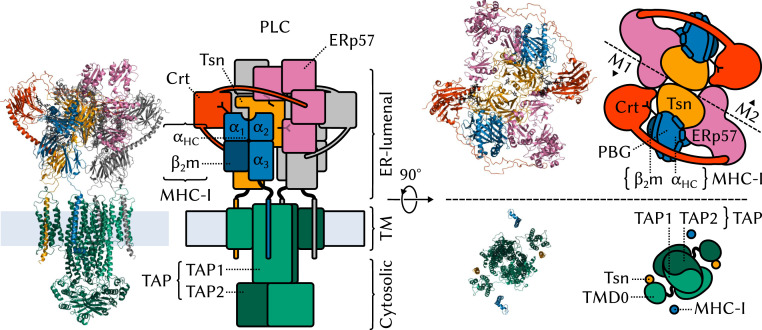
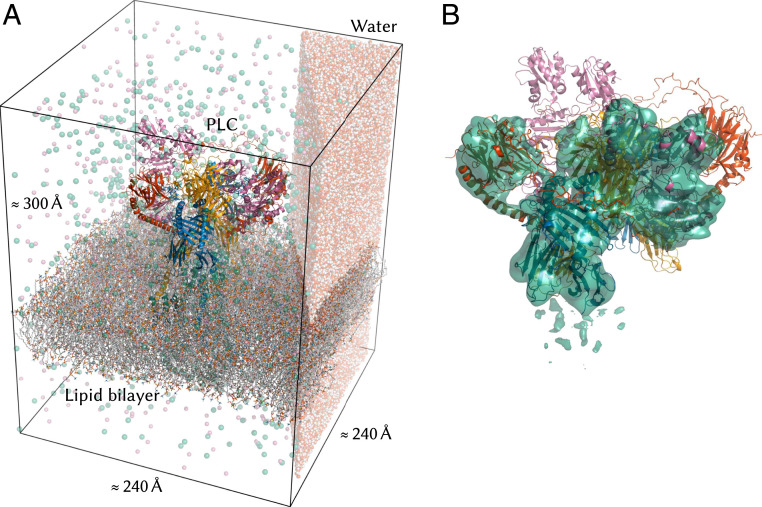
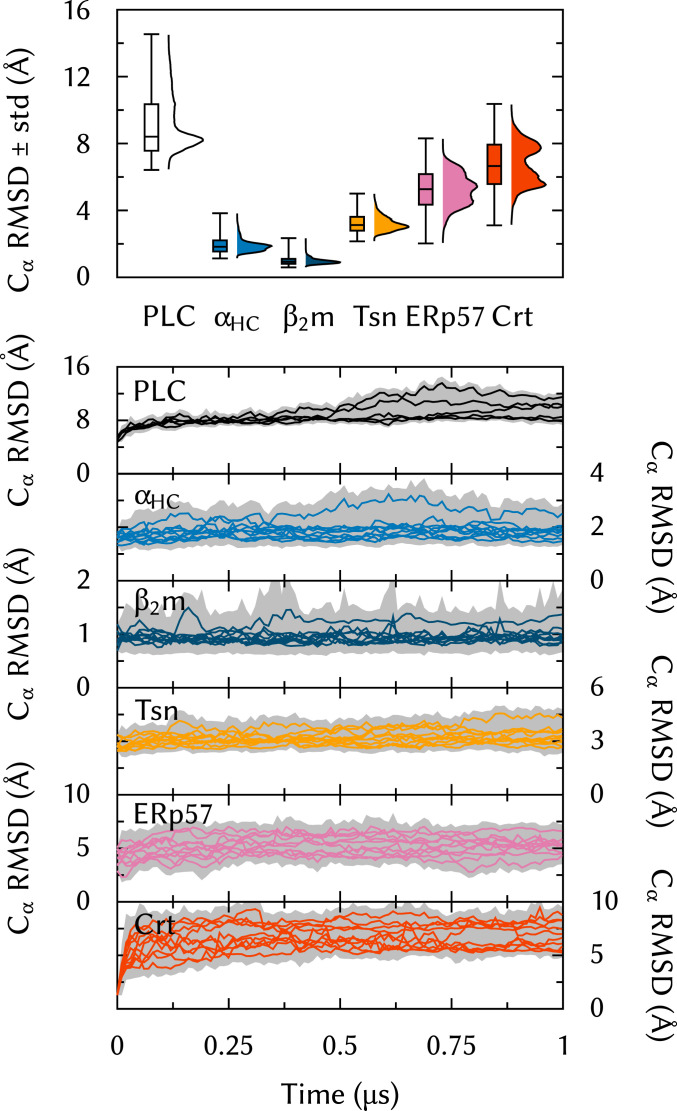
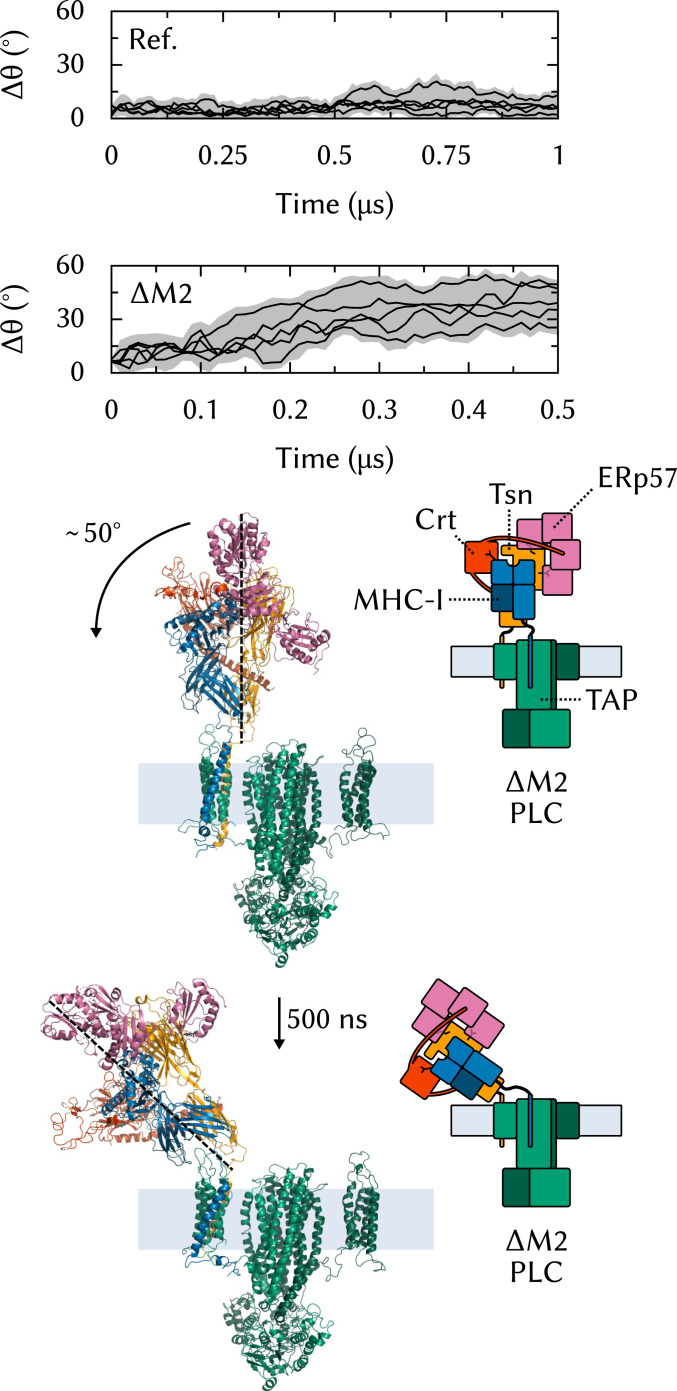
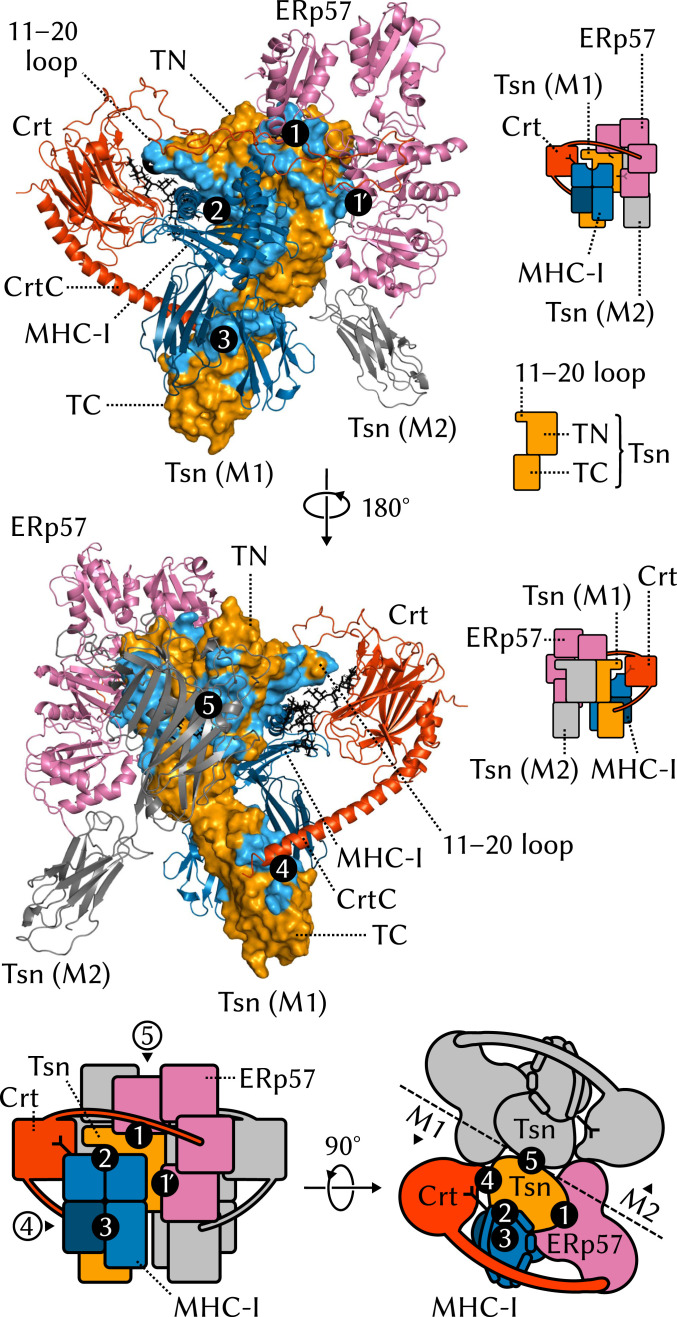
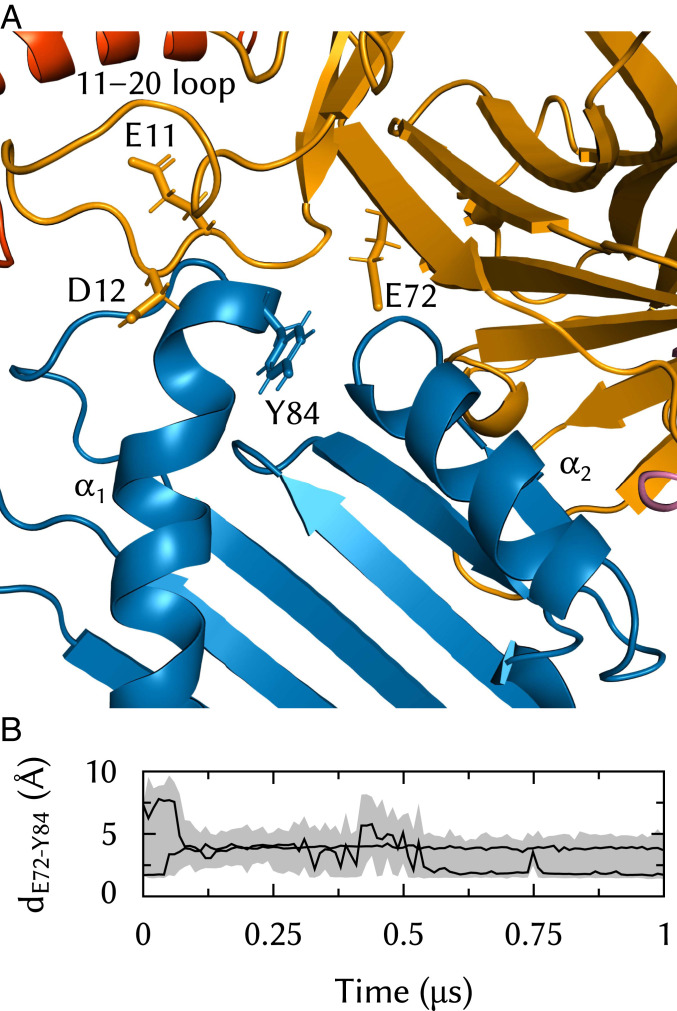
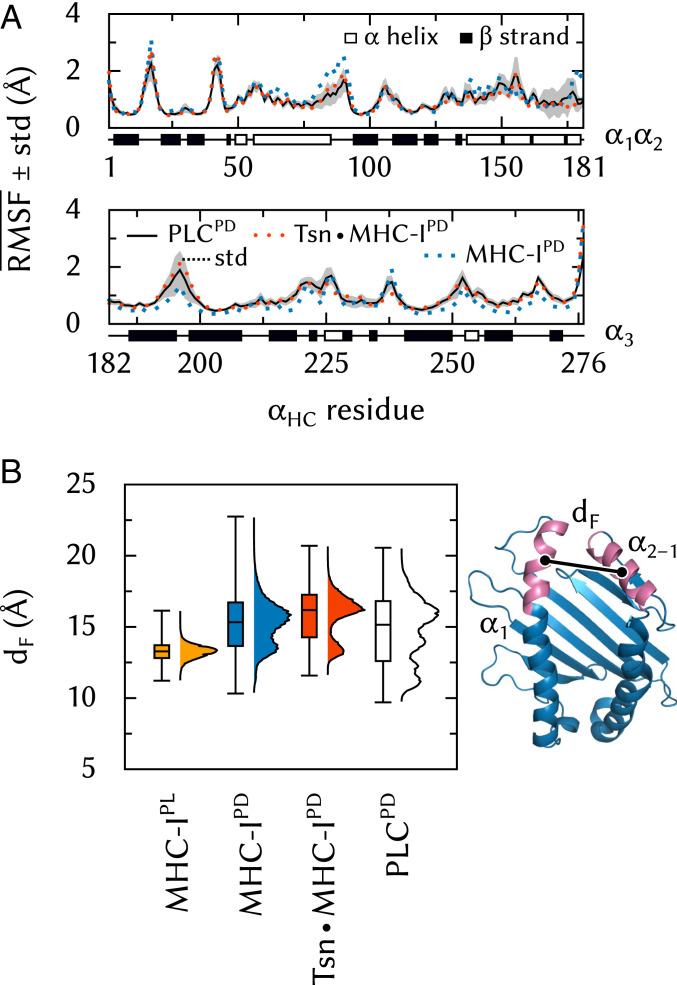
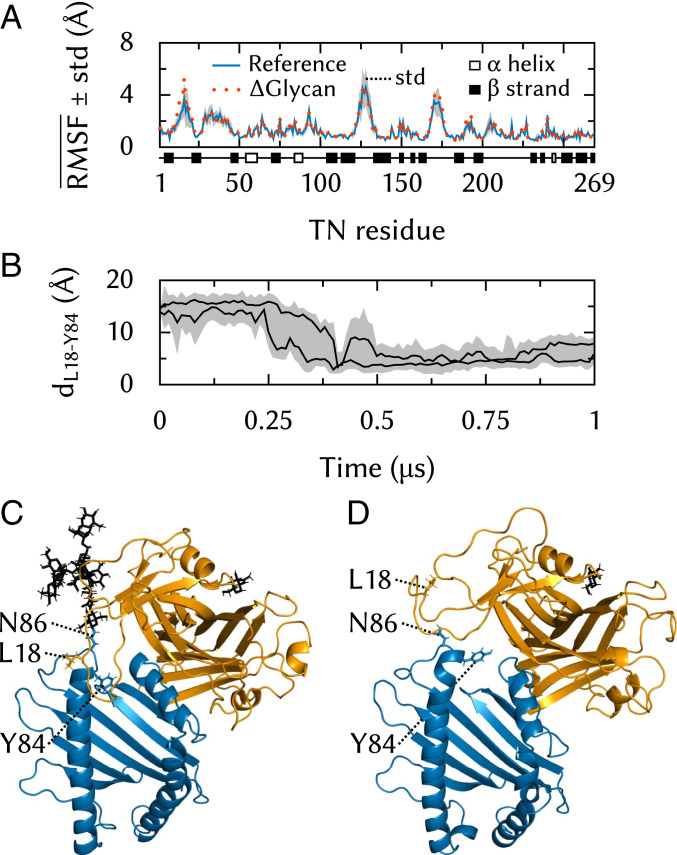
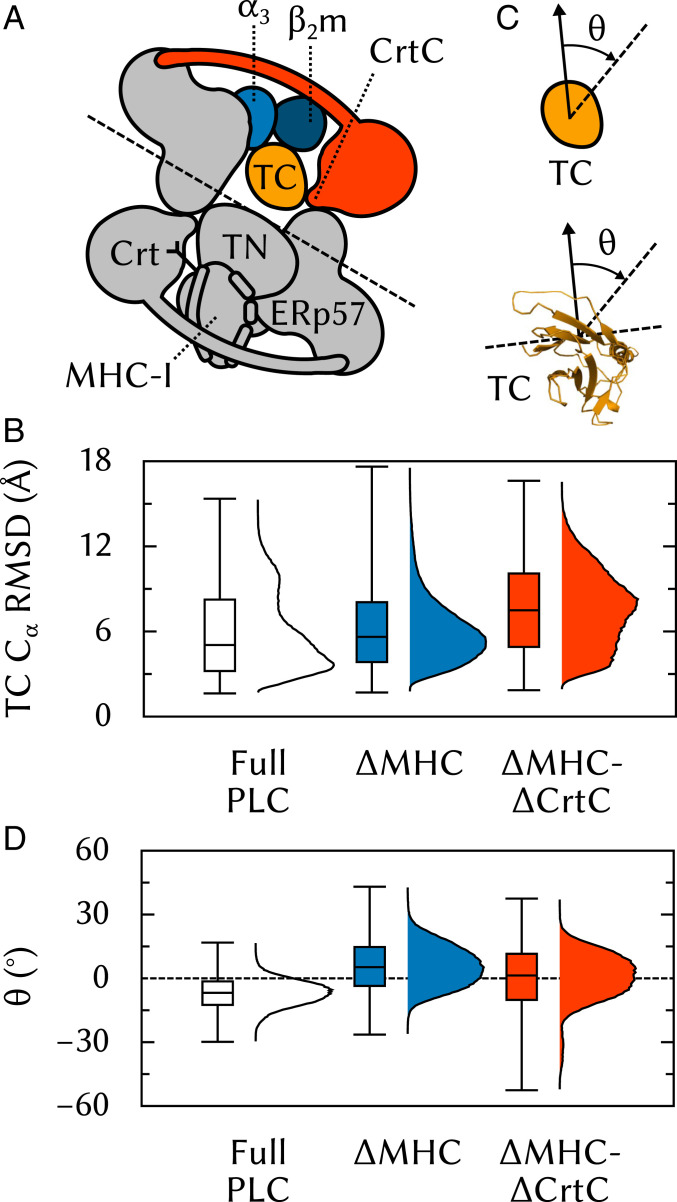
The major histocompatibility complex class-I (MHC-I) peptide-loading complex (PLC) is a cornerstone of the human adaptive immune system, being responsible for processing antigens that allow killer T cells to distinguish between healthy and compromised cells. Based on a recent low-resolution cryo-electron microscopy (cryo-EM) structure of this large membrane-bound protein complex, we report an atomistic model of the PLC and study its conformational dynamics on the multimicrosecond time scale using all-atom molecular dynamics (MD) simulations in an explicit lipid bilayer and water environment (1.6 million atoms in total). The PLC has a layered structure, with two editing modules forming a flexible protein belt surrounding a stable, catalytically active core. Tapasin plays a central role in the PLC, stabilizing the MHC-I binding groove in a conformation reminiscent of antigen-loaded MHC-I. The MHC-I-linked glycan steers a tapasin loop involved in peptide editing toward the binding groove. Tapasin conformational dynamics are also affected by calreticulin through a conformational selection mechanism that facilitates MHC-I recruitment into the complex.
Keywords: MHC-I; immunity; molecular dynamics simulations; peptide-loading complex; protein dynamics.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Neefjes J., Jongsma M. L. M., Paul P., Bakke O., Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 11, 823–836 (2011). - PubMed
-
- Hulpke S., Tampé R., The MHC I loading complex: A multitasking machinery in adaptive immunity. Trends Biochem. Sci. 38, 412–420 (2013). - PubMed
-
- Blees A., et al. , Structure of the human MHC-I peptide-loading complex. Nature 551, 525–528 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
