

Molecular and cellular basis of left-right asymmetry in vertebrates
- PMID: 32788551
- PMCID: PMC7443379
- DOI: 10.2183/pjab.96.021
Molecular and cellular basis of left-right asymmetry in vertebrates
Abstract
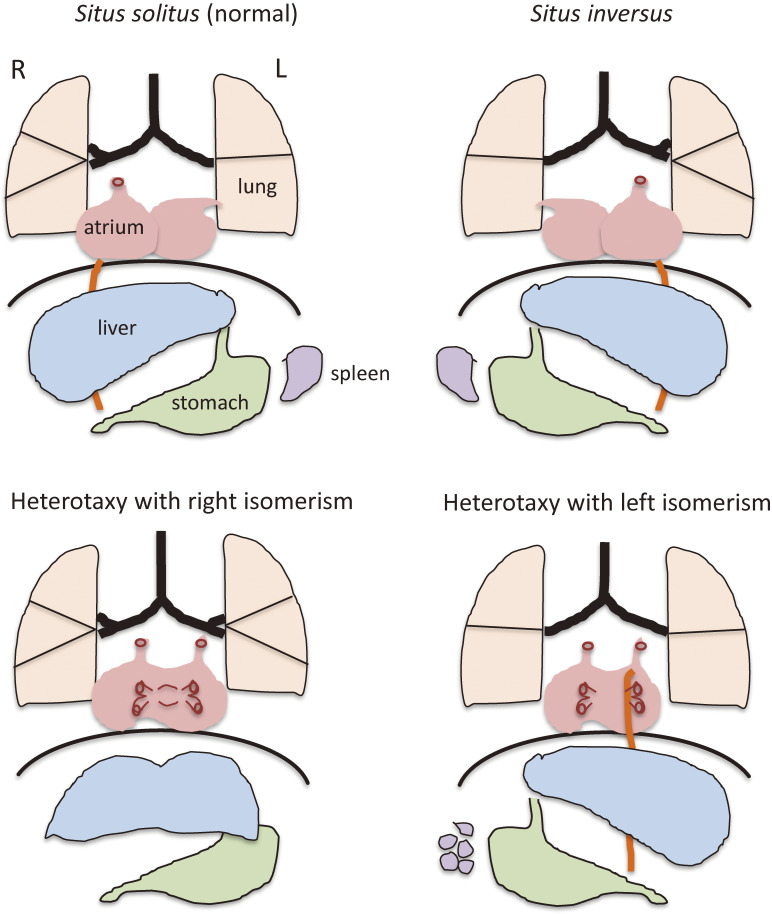
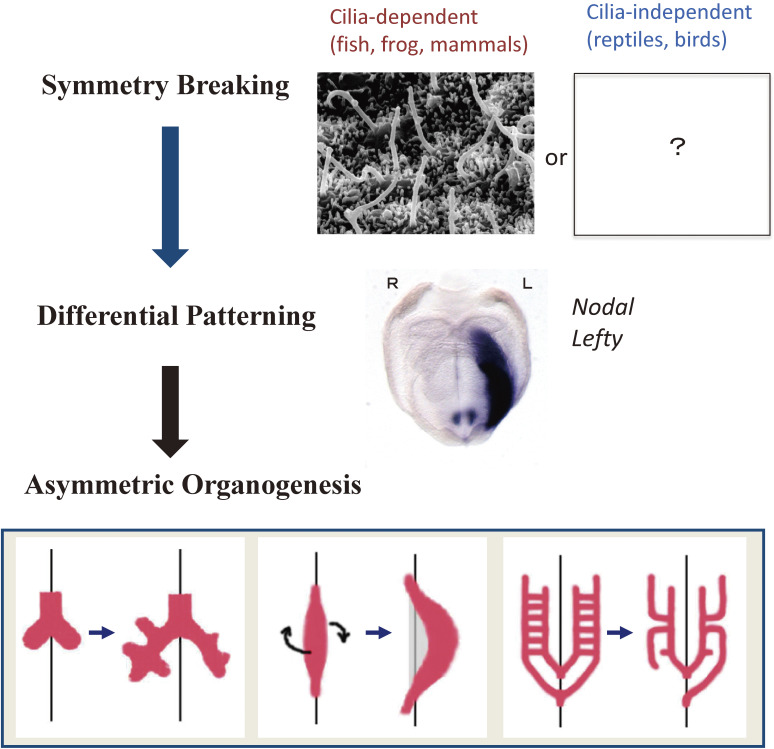
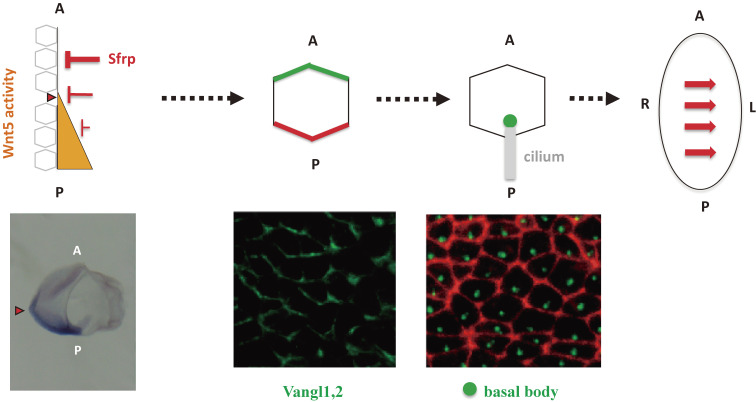
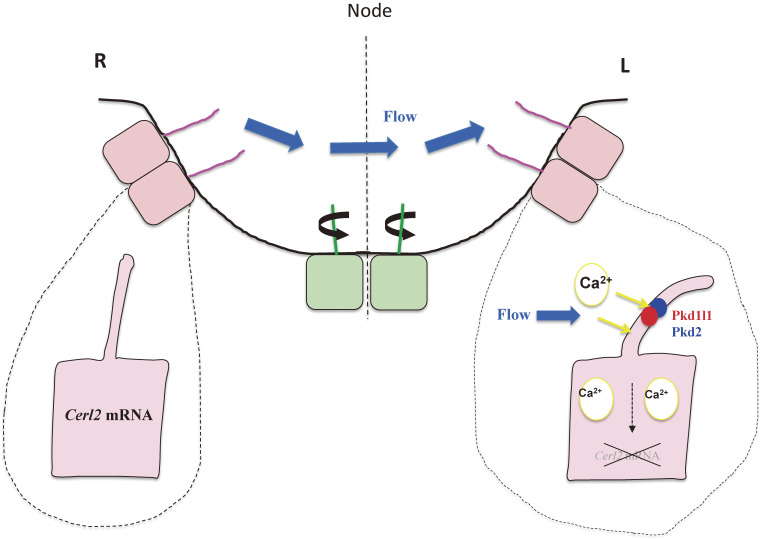
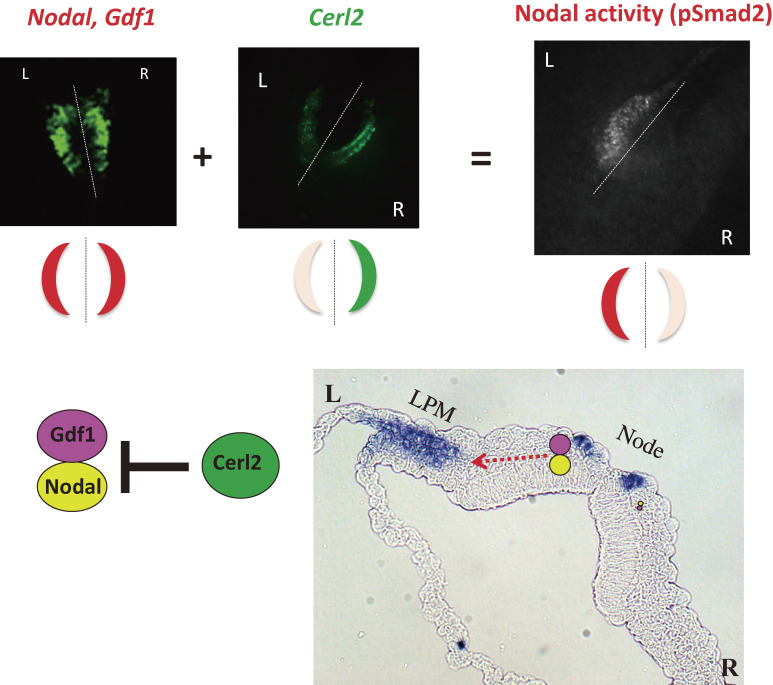
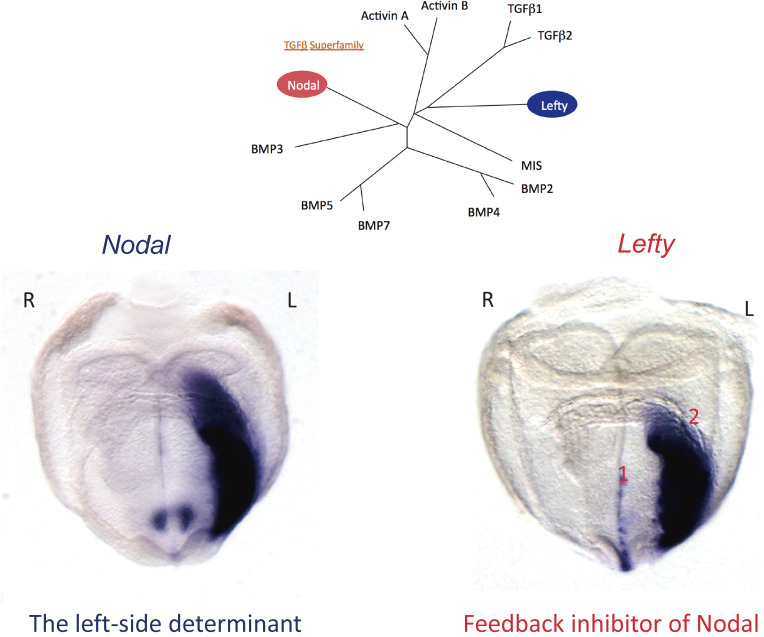
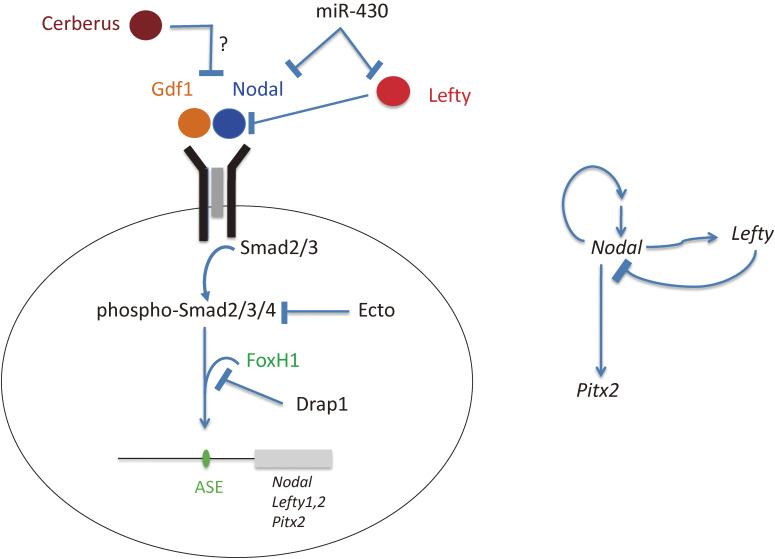
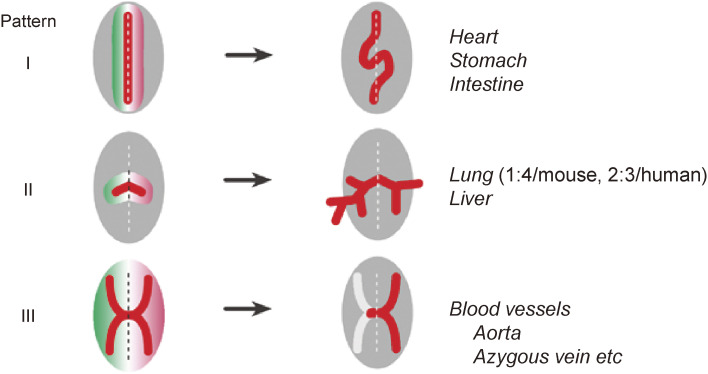
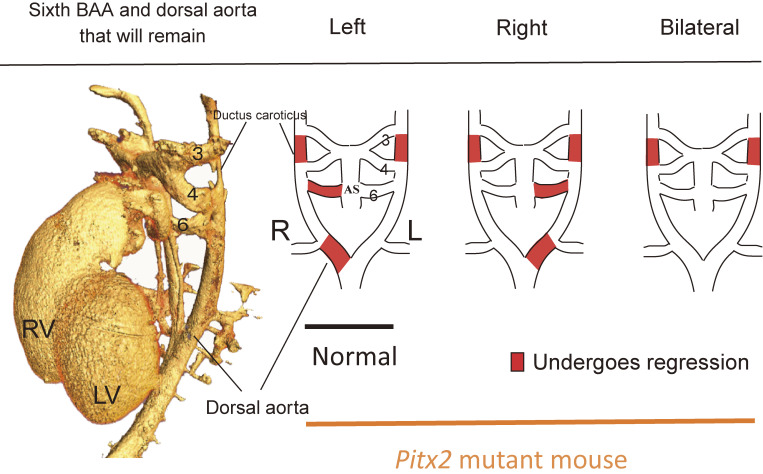
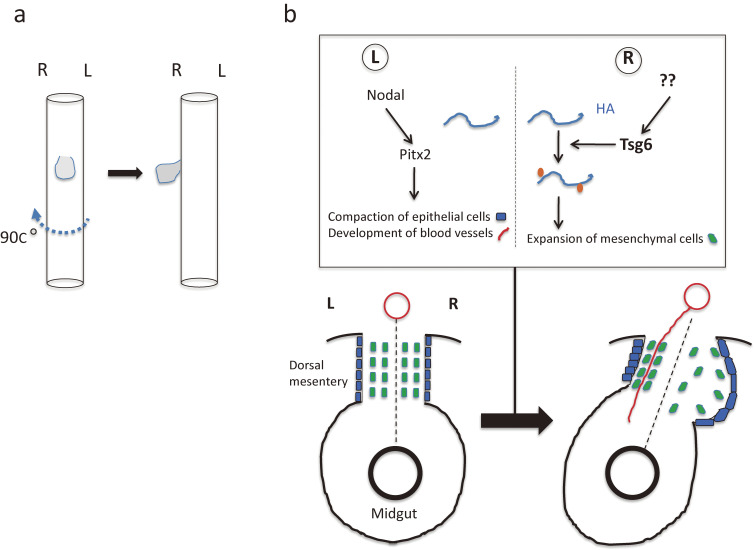
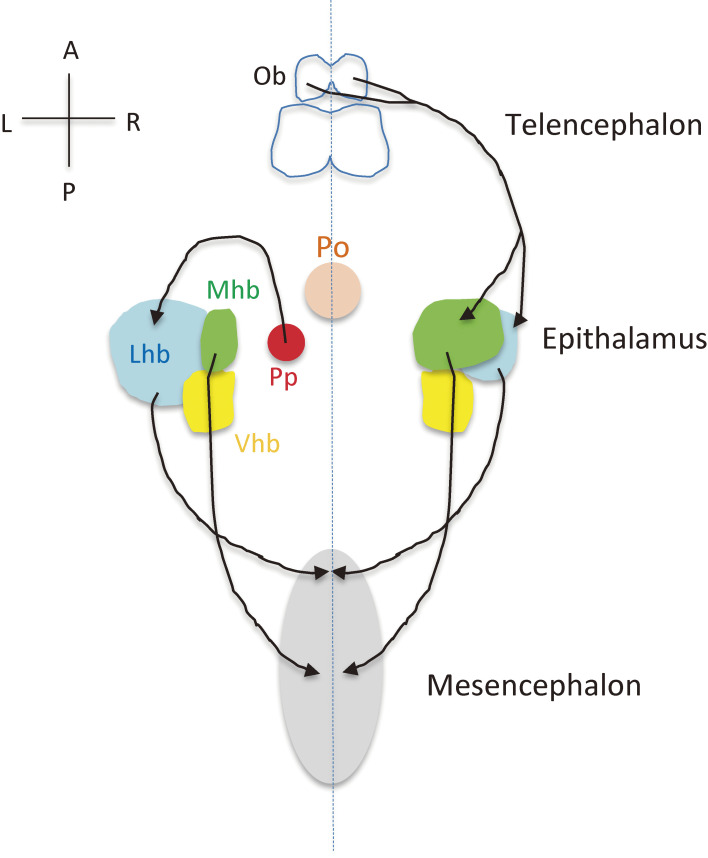
Although the human body appears superficially symmetrical with regard to the left-right (L-R) axis, most visceral organs are asymmetric in terms of their size, shape, or position. Such morphological asymmetries of visceral organs, which are essential for their proper function, are under the control of a genetic pathway that operates in the developing embryo. In many vertebrates including mammals, the breaking of L-R symmetry occurs at a structure known as the L-R organizer (LRO) located at the midline of the developing embryo. This symmetry breaking is followed by transfer of an active form of the signaling molecule Nodal from the LRO to the lateral plate mesoderm (LPM) on the left side, which results in asymmetric expression of Nodal (a left-side determinant) in the left LPM. Finally, L-R asymmetric morphogenesis of visceral organs is induced by Nodal-Pitx2 signaling. This review will describe our current understanding of the mechanisms that underlie the generation of L-R asymmetry in vertebrates, with a focus on mice.
Keywords: Nodal; chirality; cilia; left–right asymmetry.
Figures



References
-
- Smith C.A., Sinclair A.H. (2004) Sex determination: Insights from the chicken. Bioessays 26, 120–132. - PubMed
-
- Kartagener M. (1933) Zur pathogenese der bronchiektasen. 1. Mitteilung: Bronchiektasien bei situs viscerum inversus. Beitr. Klin. Tuberk. 83, 489–501.
-
- Hummel K.P. (1959) Developmental anomalies in mice resulting from action of the gene, disorganization, a semi-dominant lethal. Pediatrics 23, 212–221. - PubMed