

Serine restriction alters sphingolipid diversity to constrain tumour growth
- PMID: 32788725
- PMCID: PMC7606299
- DOI: 10.1038/s41586-020-2609-x
Serine restriction alters sphingolipid diversity to constrain tumour growth
Abstract
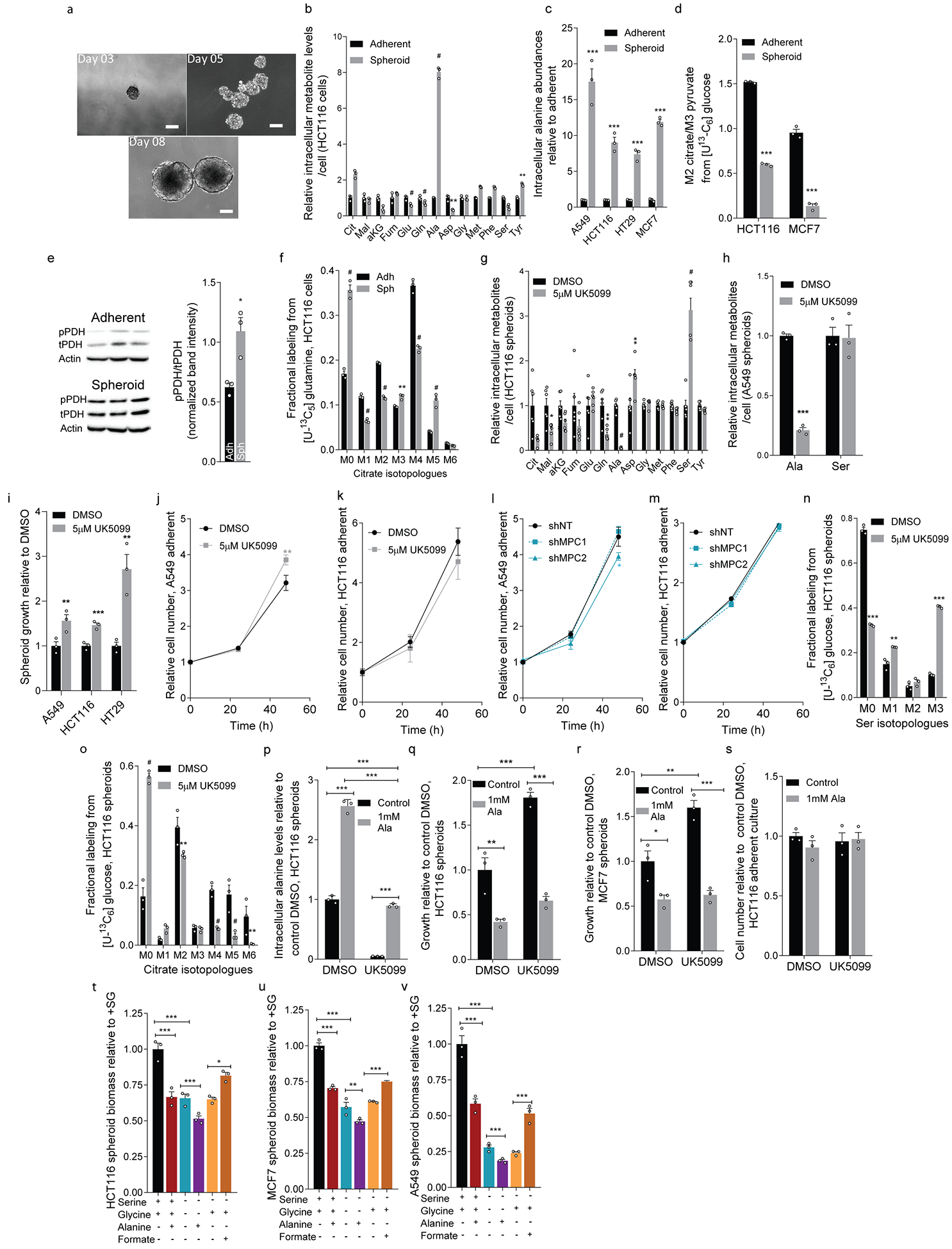
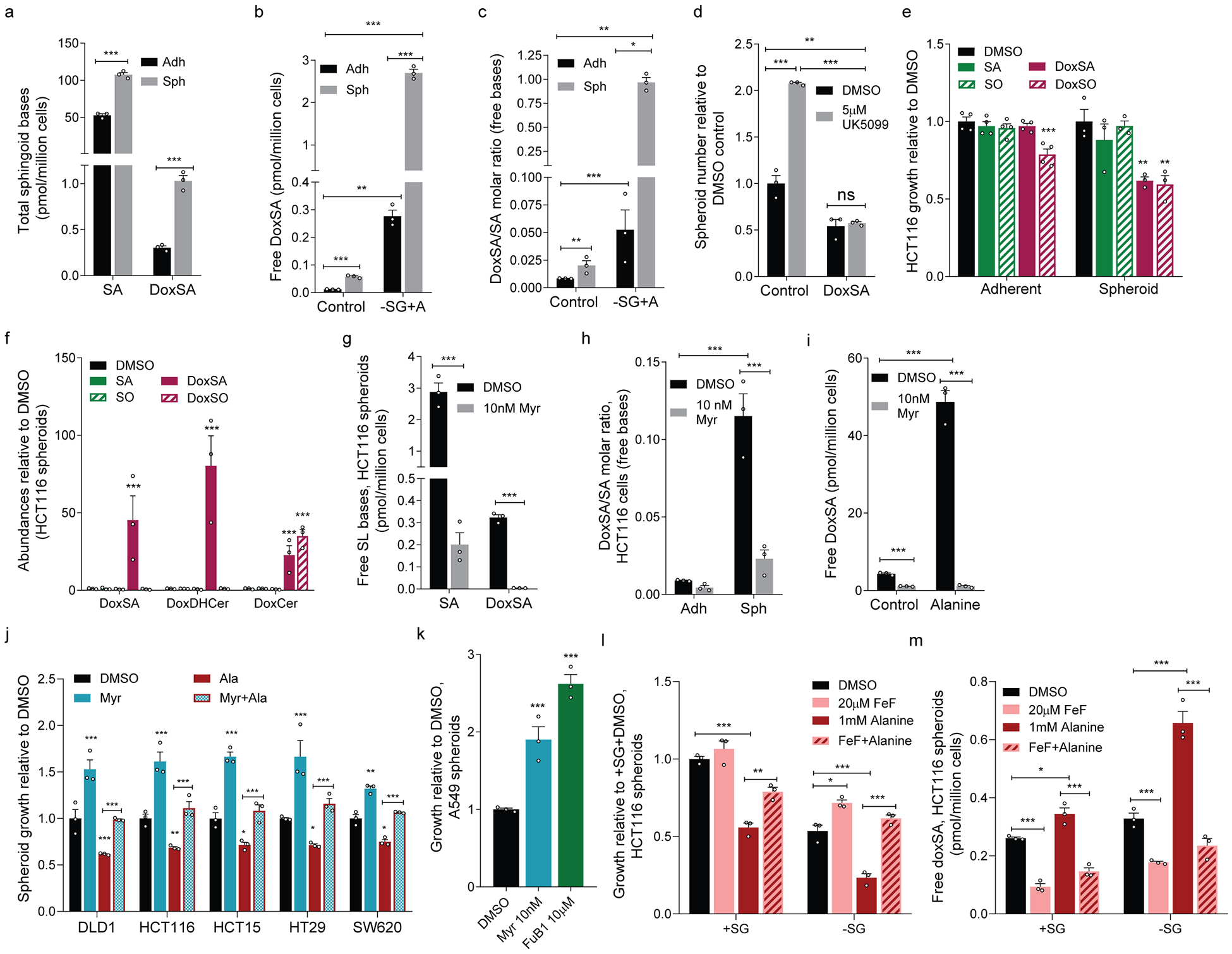
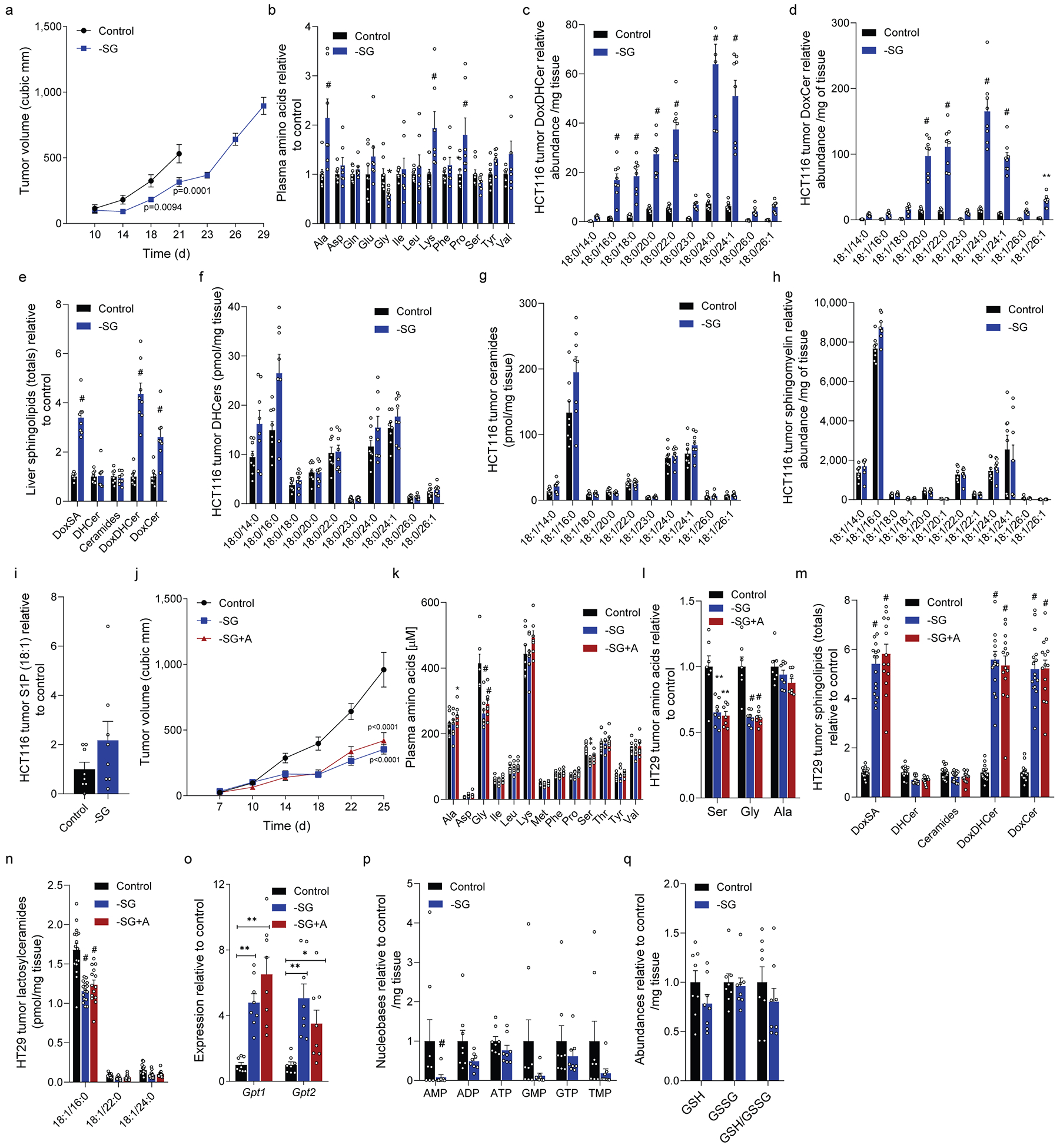
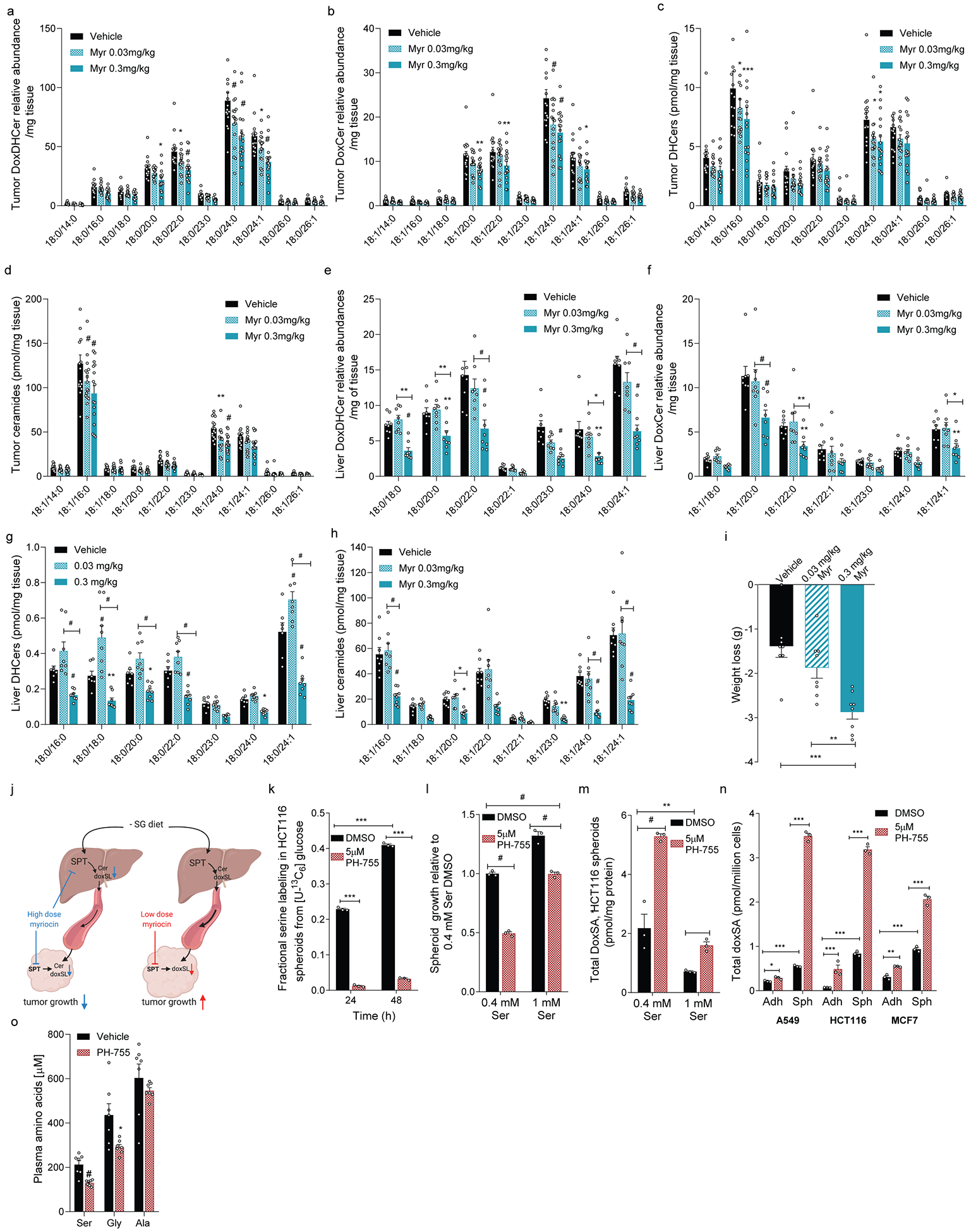
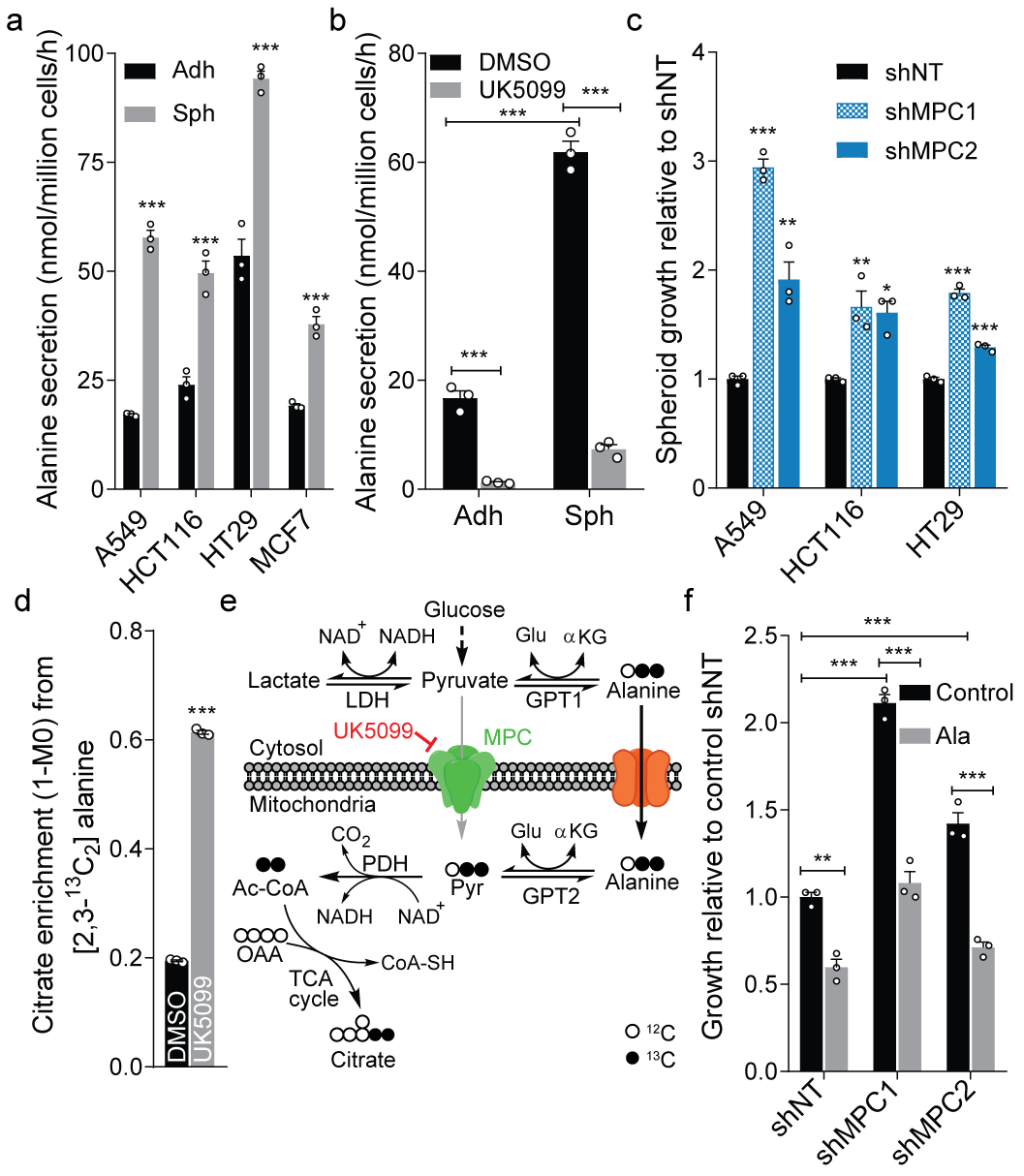
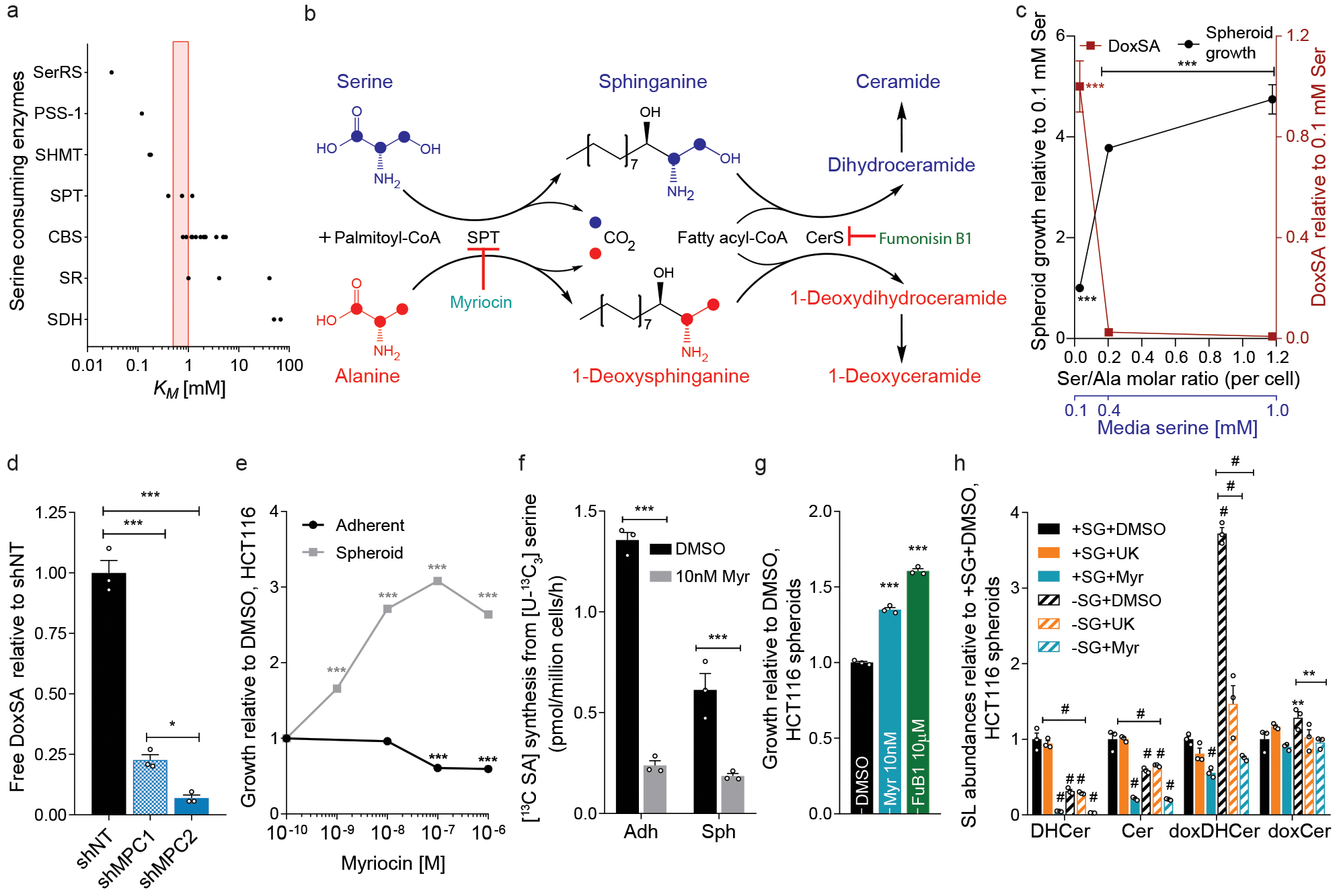
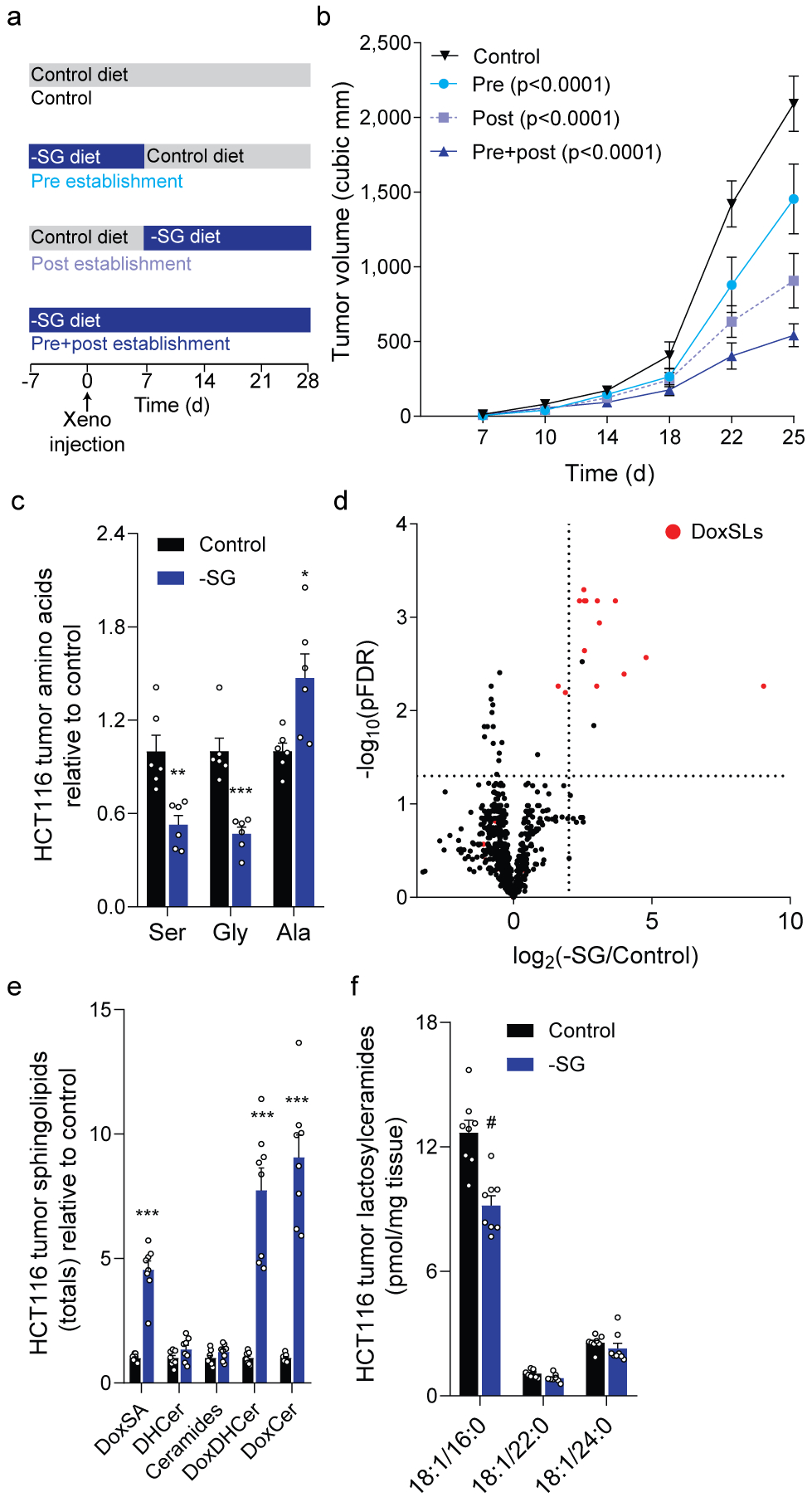
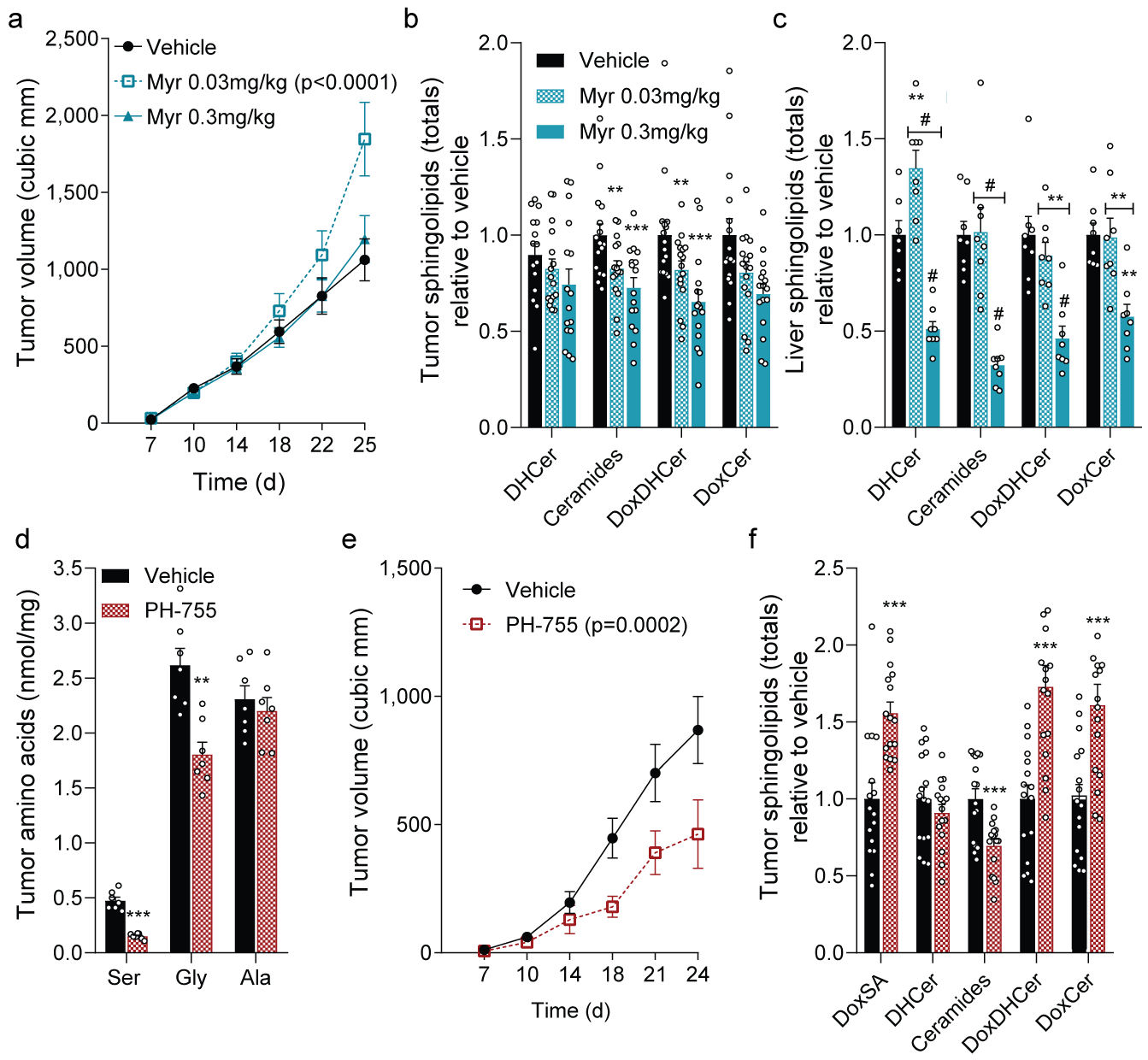
Serine, glycine and other nonessential amino acids are critical for tumour progression, and strategies to limit their availability are emerging as potential therapies for cancer1-3. However, the molecular mechanisms driving this response remain unclear and the effects on lipid metabolism are relatively unexplored. Serine palmitoyltransferase (SPT) catalyses the de novo biosynthesis of sphingolipids but also produces noncanonical 1-deoxysphingolipids when using alanine as a substrate4,5. Deoxysphingolipids accumulate in the context of mutations in SPTLC1 or SPTLC26,7-or in conditions of low serine availability8,9-to drive neuropathy, and deoxysphinganine has previously been investigated as an anti-cancer agent10. Here we exploit amino acid metabolism and the promiscuity of SPT to modulate the endogenous synthesis of toxic deoxysphingolipids and slow tumour progression. Anchorage-independent growth reprogrammes a metabolic network involving serine, alanine and pyruvate that drives the endogenous synthesis and accumulation of deoxysphingolipids. Targeting the mitochondrial pyruvate carrier promotes alanine oxidation to mitigate deoxysphingolipid synthesis and improve spheroid growth, similar to phenotypes observed with the direct inhibition of SPT or ceramide synthesis. Restriction of dietary serine and glycine potently induces the accumulation of deoxysphingolipids while decreasing tumour growth in xenograft models in mice. Pharmacological inhibition of SPT rescues xenograft growth in mice fed diets restricted in serine and glycine, and the reduction of circulating serine by inhibition of phosphoglycerate dehydrogenase (PHGDH) leads to the accumulation of deoxysphingolipids and mitigates tumour growth. The promiscuity of SPT therefore links serine and mitochondrial alanine metabolism to membrane lipid diversity, which further sensitizes tumours to metabolic stress.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Comment in
-
SERIneALanine Killer: SPT promiscuity inhibits tumour growth via intra-tumoral deoxysphingolipid production.Signal Transduct Target Ther. 2020 Nov 24;5(1):274. doi: 10.1038/s41392-020-00401-6. Signal Transduct Target Ther. 2020. PMID: 33235228 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
