

Comment
doi: 10.1038/s41586-020-2510-7.
Epub 2020 Aug 12.
Heat detection by the TRPM2 ion channel
Affiliations
- PMID: 32788732
- PMCID: PMC7613795
- DOI: 10.1038/s41586-020-2510-7
Item in Clipboard
Comment
Heat detection by the TRPM2 ion channel
Nature.
2020 Aug.
No abstract available
Figures
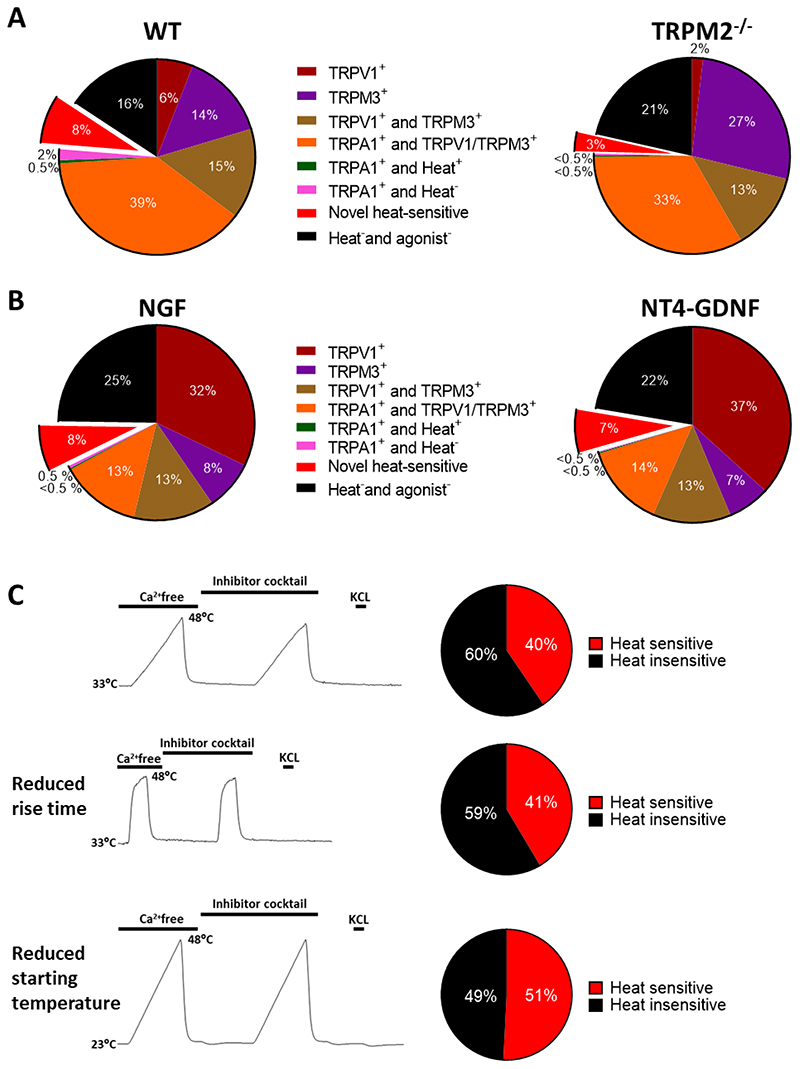
A. Neurons from trigeminal ganglion (TG) tested for heat and agonist sensitivity using protocol shown in Fig. 2A, but without application of 2-APB. Left: Percentage distributions of heat and agonist-sensitive neurons. TRPA1 is strongly co-expressed with TRPV1 or TRPM3 (39%), with few neurons expressing TRPA1 alone (2.5%). Novel heat-sensitive neurons that respond to heat but do not express any of TRPV1, TRPM3 or TRPA1 can be clearly identified (8%). n = 388 neurons from 1 WT mouse. Right: Similar experiment on n = 382 TG neurons from TRPM2-/- mouse. Proportion of novel heat sensitive neurons significantly reduced (from 8% to 3%, p = 0.0077, Fisher’s exact test). Mean response amplitude of novel heat-sensitive neurons not reduced (ΔF340/F380 = 1.13 ± 0.05, n=31, to 0.96 ± 0.08, n=13). Difference not significant, p = 0.0539, unpaired t-test. B. Neurons from DRG tested for heat and agonist sensitivity using protocol shown in Fig. 2A, but without application of 2-APB and with agonists presented after heat. Left: Neurons cultured for 12h in NGF. See Methods for details. Pie chart shows data from 3 experiments on a total of 1312 neurons from 3 WT male mice on 12 coverslips as follows: neurons expressing only TRPV1 (32.2 ± 4.2%, mean ± SEM); only TRPM3 (8.2 ± 1.2%); both TRPV1 and TRPM3 but not TRPA1 (13.3 ± 0.6%); TRPA1 together with either TRPV1 or TRPM3 (13.3 ± 0.9%); TRPA1 alone, and responding to heat (0.2 ± 0.2%); TRPA1 alone and not responding to heat (0.4 ± 0.2%); novel heat-sensitive neurons responding to heat but not to agonists for any of TRPV1, TRPM3 or TRPA1 (7.6 ± 2.0%); and neurons not responding to heat nor to any of the TRP agonists but identified as viable from response to final pulse of KCl (24.8 ± 1.8%). The application of agonists before (Fig. 1) or after heat pulse (this Fig) does not significantly affect the proportion of novel heat-sensitive neurons detected (9.5 ± 0.2% when agonists applied before, see Fig. 1A, and 7.6 ± 2.0% when after (p = 0.3740, unpaired t-test)). Proportions of neurons in other categories also not significantly affected, apart from a small reduction in the proportion of neurons expressing TRPA1 in combination with TRPV1 and/or TRPM3 (compare Fig. 1A). Right: Neurons cultured for 12h in NT4-GDNF. n = 1521 neurons from 3 WT male mice on 12 coverslips. These culture conditions had no significant effect on the proportions of heat and agonist-sensitive neurons. p>0.05, multiple t test with Bonferroni correction. C. Protocol for determining total proportion of novel heat-sensitive neurons from DRG, including those co-expressing TRPV1, TRPM3 and/or TRPA1. Heat ramp is first applied in Ca-free solution, to identify changes in F340/F380 fluorescence ratio (ΔF340/F380) arising from sensitivity of the fura2 dye to temperature, followed by an identical ramp in the presence of a cocktail of inhibitors of TRPV1 (AMG9810, 5μM), TRPM3 (naringenin, 10μM), and TRPA1 (HC-030031, 100μM) (see details in Methods). Final pulse of KCl confirms neuronal viability. Top: Heat application rising from 33°C to 48°C in 180 sec1. n = 710 neurons from 2 WT mice on 3 coverslips. Middle: Heat application rising from 33°C to 48°C in 25 sec2. n= 99 neurons from 1 WT mouse on 2 coverslips. Bottom: Heat application rising from 23°C to 48°C in 180 sec2. n= 657 neurons from 1 WT mouse on 2 coverslips. Reducing rise time or starting temperature of heat application does not reduce the total proportion of novel heat-sensitive neurons. Difference not significant, p = 0.9197 (middle) and 0.3339 (bottom) respectively, unpaired t-test compared to top panel.
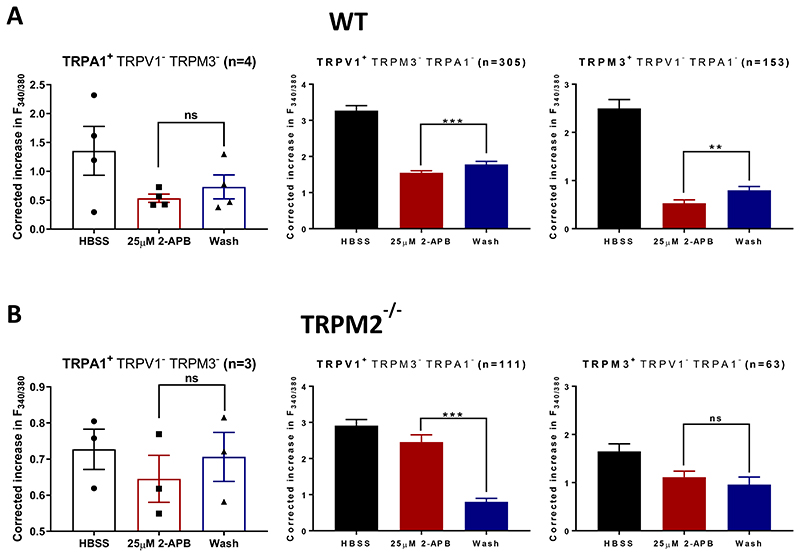
Neurons were identified as expressing only one of TRPA1 (left), TRPV1 (middle) or TRPM3 (right) and the effect of 2-APB (25μM) was tested as shown in Fig. 2A. A, WT neurons. Note that many of these neurons will co-express TRPM2, which is extensively expressed in somatosensory neurons (Fig. 1C). B, Similar experiments on TRPM2-/- neurons. Significance level during 2-APB application compared to that after wash is shown as: *, p < 0.05; **, p < 0.01; ***, p < 0.001; n.s., p > 0.05. One-way ANOVA followed by Bonferroni’s post-hoc correction.
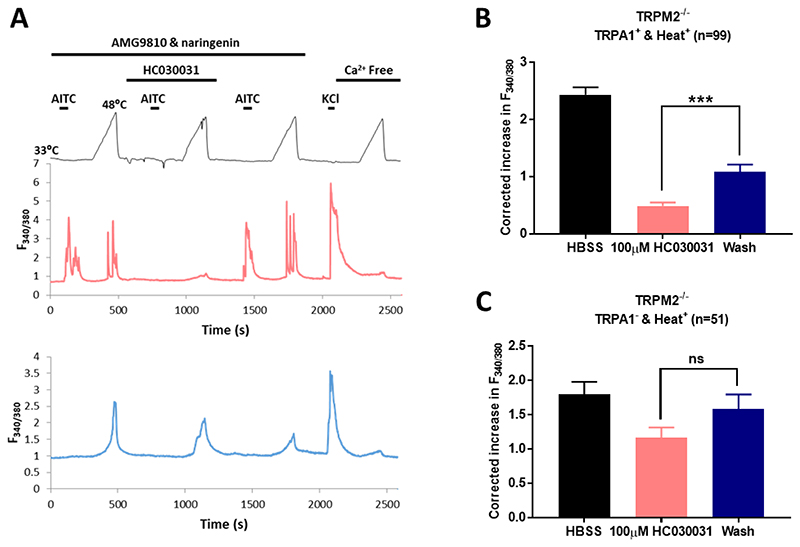
A. Upper: Protocol of solution changes used to identify neurons expressing TRPA1 and to test their activation by heat. TRPV1 and TRPM3 were blocked by AMG9810 (5μM) and naringenin (10μM) as shown. The calcium increase elicited by a heat ramp from 33°C to 48°C was tested in the absence (heat ramps 1 and 3) and presence (ramp 2) of the TRPA1 blocker HC030031 (100μM). Responses to the TRPA1-selective agonist AITC were tested as shown. Response to KCl (50mM) and heat ramp in absence of external Ca tested at end. Middle trace: increases in F340/380 in a neuron expressing TRPA1 in which the heat-activated Ca increase was suppressed by the TRPA1 blocker HC030031. TRPA1 was expressed in 31.2 ± 4.2% of the total population, but only a fraction of TRPA1+ neurons (34.7% ± 9.81%) gave a significant Ca increase in response to 48°C heat. Lower trace: a different neuron in same preparation is activated by heat but does not express TRPA1 (unidentified heat-responders). Data from 3 separate experiments on a total of 964 neurons. B. Heat response in TRPA1-expressing neurons is suppressed by TRPA1 blocker HC030031 (protocol and example trace shown in A. ***, p<0.001, one-way ANOVA followed by Bonferroni’s post-hoc correction). Heat response in neurons not expressing TRPA1 is not significantly reduced by the TRPA1 blocker HC030031 (100μM). Proportion of unidentified heat-responders (5.5% ± 1.4%) is not significantly different from that in Fig. 1D.
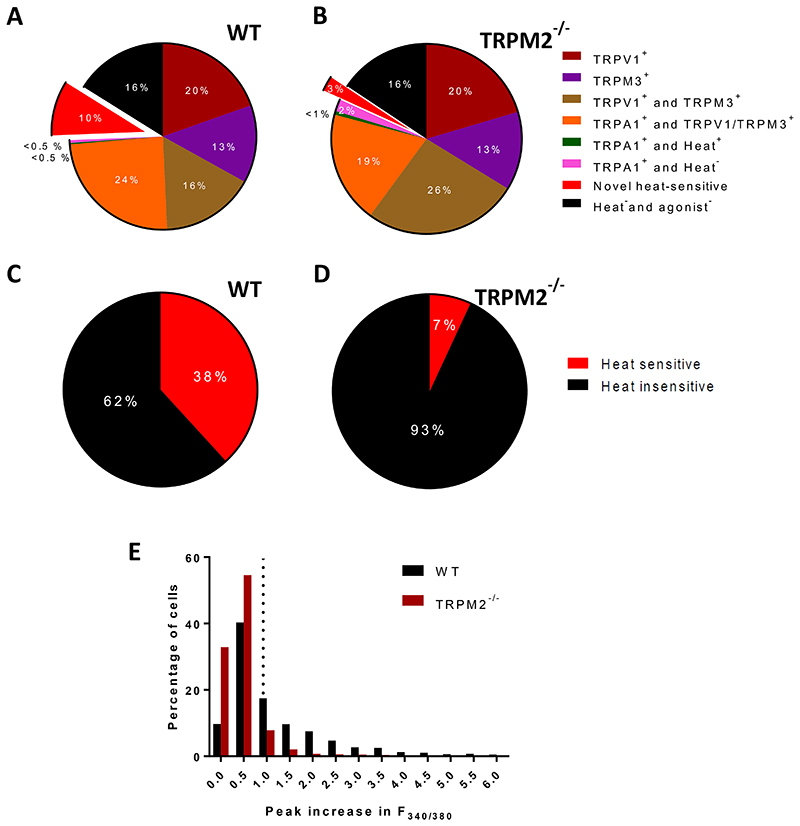
A. Proportions of DRG neurons expressing TRPV1, TRPM3 and TRPA1, and of novel heat sensitive neurons expressing none of these TRP channels, from increases in F340/F380 in response to application of selective agonists (see Methods for details). Experimental protocol shown in Fig. 2A (except that TRPM2 blocker 2-APB was not applied). Pie chart shows data from 3 experiments as follows: neurons expressing only TRPV1 (19.6 ± 2.0%, mean ± SEM); only TRPM3 (13.5 ± 0.2%); both TRPV1 and TRPM3 but not TRPA1 (16.2 ± 3.5%); TRPA1 together with either TRPV1 or TRPM3 (24.4 ± 1.6%); TRPA1 alone, and responding to heat (0.2 ± 0.1%); TRPA1 alone and not responding to heat (0.49 ± 0.2%); novel heat-sensitive neurons responding to heat but not to agonists for any of TRPV1, TRPM3 or TRPA1 (9.5 ± 0.2%); and neurons not responding to heat nor to any of the TRP agonists but identified as viable from response to final pulse of KCl (16.1 ± 2.6%). See criteria in Methods for identifying positive heat and agonist responses. Data from 3 separate experiments on a total of 1563 neurons (13 cover slips from 3 male wild-type mice). B. Similar experiment performed on DRG neurons from TRPM2-/- mice. There is no significant difference in the proportions of any of the agonist-sensitive neurons from WT (p>0.05, multiple t test with Bonferroni correction). However, the proportion of novel heat-sensitive neurons is significantly reduced by deletion of TRPM2, from 9.5 ± 0.2% in WT to 2.5 ± 0.8% in TRPM2-/-(p = 0.009, multiple t test with Bonferroni correction). Data from 3 separate experiments on a total of 660 neurons (14 cover slips from 3 male TRPM2-/- mice). C. Proportion of DRG neurons responding to heat in the presence of inhibitor cocktail blocking TRPV1, TRPM3 and TRPA1. See Extended data Fig. 1C, top trace, for protocol. In 4 separate experiments percentage of heat-responsive neurons = 38.2 ± 4.4%, and Δ F340/F380 = 1.90 ± 0.06 (mean ± SEM). Results from 1315 WT neurons. D. Similar experiment to that shown in C, on DRG neurons from TRPM2-/- mouse. Mean percentage of “unidentified” heat-responsive neurons in n = 4 separate experiments was 6.9 ± 2.2% (mean ± SEM, significant difference from WT, p = 0.0007, unpaired t-test) and ΔF340/F380 = 1.59 ± 0.16 (mean ± SEM, significant difference from WT, p = 0.0237, unpaired t-test). Results from 1794 neurons. E. Amplitudes of ΔF340/F380 in response to heat ramp to 48°C from the experiments shown in C (WT neurons) and D (TRPM2-/- neurons). Threshold for accepting a heat response shown by vertical dotted line giving intrinsic heat-sensitivity of the fura2 dye obtained from heat ramp applied in Ca-free solution (see details in Methods). Mean ΔF340/F380 in response to heat (48°C) in all WT neurons (i.e. both heat-sensitive and heat-insensitive) was 1.24 ± 0.04 (n = 1315) and in all TRPM2-/- neurons was 0.46 ± 0.01 (n = 1794). Difference significant, p <0.0001, unpaired t-test.
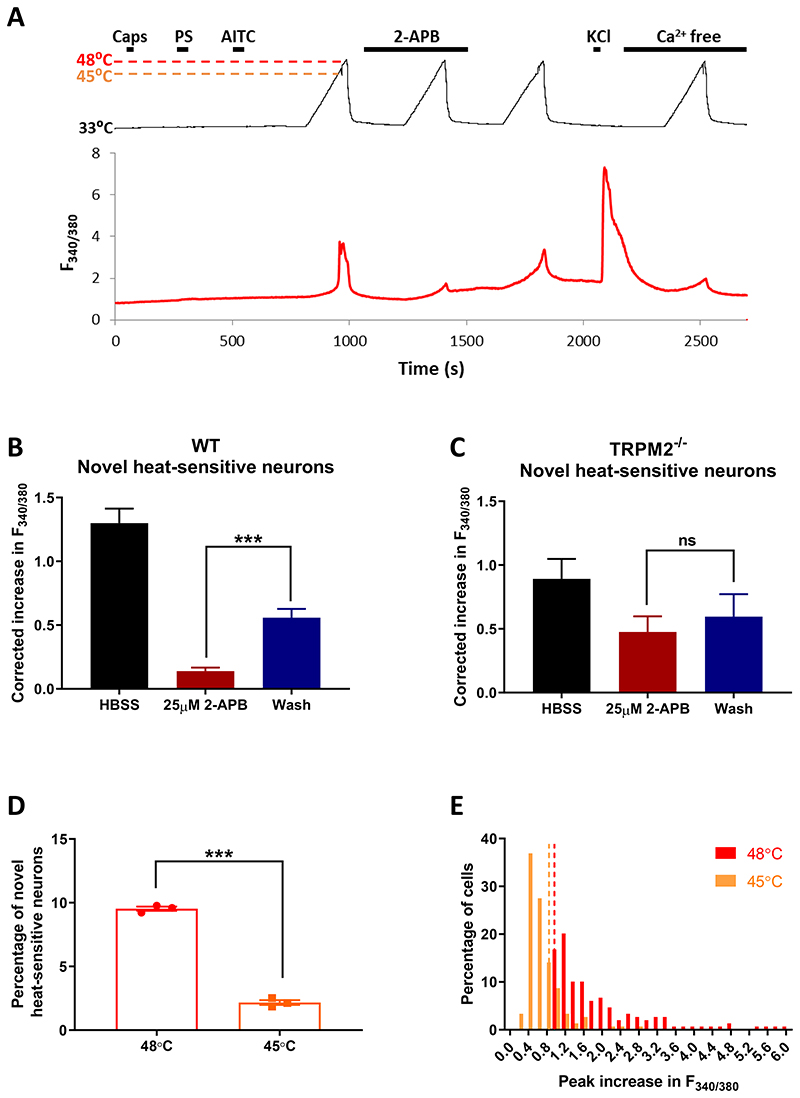
A. Effect of TRPM2 blocker 2-APB (25μM) on identified somatosensory neurons. Neurons expressing TRPV1, TRPM3 and/or TRPA1 identified by brief application of the specific agonists capsaicin (1μM), pregnenolone sulphate (PS, 100μM) and allyl isothiocyanate (AITC, 50 μM), respectively, followed by heat ramp to 48°C to activate all heat-sensitive neurons. Effect of TRPM2 blocker 2-aminoethoxydiphenyl borate (2-APB, 25μM) on heat-activated neurons is tested, followed by a pulse of KCl (50mM) to test neuronal viability. At end, heat pulse repeated in Ca-free solution, to identify changes in F340/F380 fluorescence ratio (ΔF340/F380) arising from sensitivity of the fura2 dye to temperature. Response to heat in a novel heat-sensitive neuron not expressing any of TRPV1, TRPM3 or TRPA1 is strongly suppressed by the TRPM2 blocker 2-APB (25μM). B. Increase in calcium-dependent fluorescence ratio (ΔF340/F380) in response to heat in novel heat-sensitive neurons (n = 149 neurons from 3 wild-type mice) is strongly suppressed by 2-APB (25μM). Increase in ΔF340/F380 corrected for each neuron by subtracting the increase in ΔF340/F380 measured from heat ramp applied in Ca-free solution. C. Similar experiment on neurons from 3 TRPM2-/- mice. The few novel-heat-sensitive neurons observed (see Fig. 1B) show no significant effect of 2-APB on response amplitude. D. Effect of terminating temperature ramp at 48°C and at an earlier time point when temperature had reached 45°C. Reducing final temperature had a dramatic effect on proportion of novel heat-sensitive neurons, from 9.5 ± 0.2% in response to heat to 48°C to 2.2 ± 0.2% in response to heat to 45°C (p<0.0001, unpaired t-test). n = 1563 neurons from 3 WT mice. E. Amplitudes of ΔF340/F380 in response to heat ramp reaching 48°C and 45°C from the experiment shown in B. Threshold for accepting a heat response shown by vertical dotted lines showing intrinsic heat-sensitivity of the fura2 dye obtained from heat ramp applied in 0Ca solution (see details in Methods). Mean ΔF340/F380 in response to heat to 48°C was 2.04 ± 0.11 and to 45°C was 0.67± 0.03 (n = 149 neurons, p<0.0001, unpaired t-test).
Comment in
-
Reply to: Heat detection by the TRPM2 ion channel.Nature. 2020 Aug;584(7820):E13-E15. doi: 10.1038/s41586-020-2511-6. Nature. 2020. PMID: 32788731 No abstract available.
Comment on
-
A TRP channel trio mediates acute noxious heat sensing.Nature. 2018 Mar 29;555(7698):662-666. doi: 10.1038/nature26137. Epub 2018 Mar 14. Nature. 2018. PMID: 29539642
References
-
- Bautista DM, et al. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell. 2006;124:1269–1282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
