

Essential Role for CD30-Transglutaminase 2 Axis in Memory Th1 and Th17 Cell Generation
- PMID: 32793209
- PMCID: PMC7385138
- DOI: 10.3389/fimmu.2020.01536
Essential Role for CD30-Transglutaminase 2 Axis in Memory Th1 and Th17 Cell Generation
Abstract
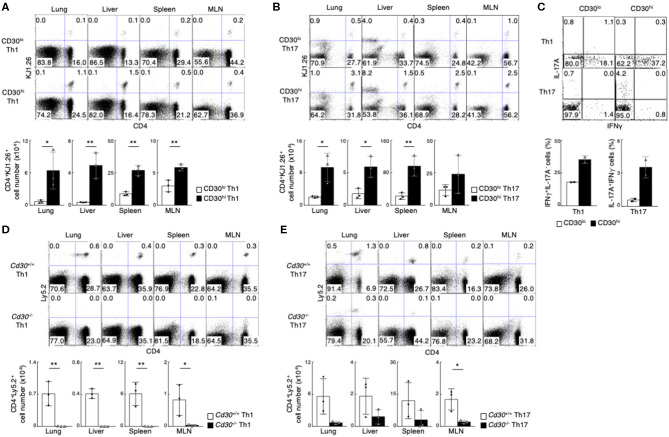
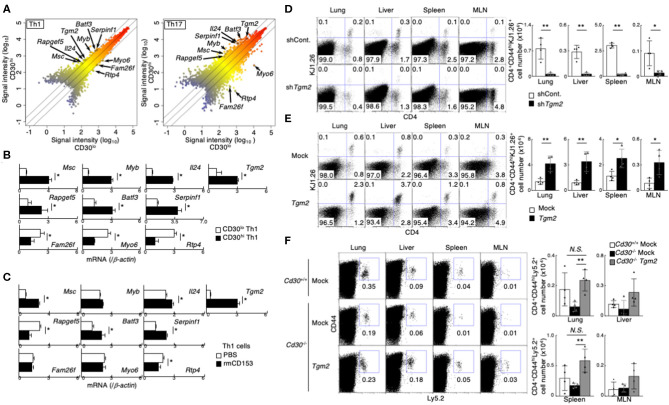
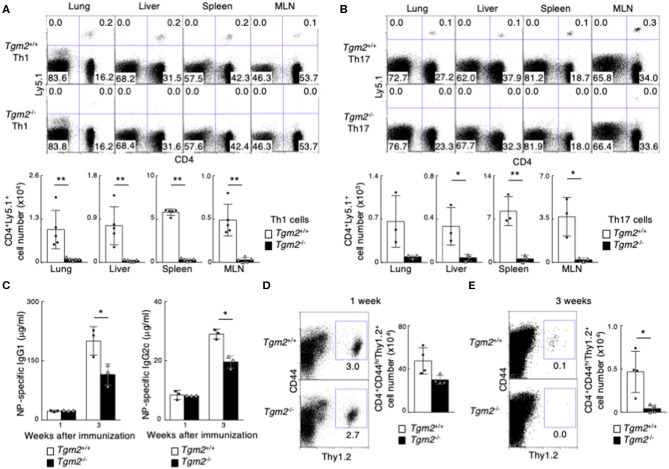
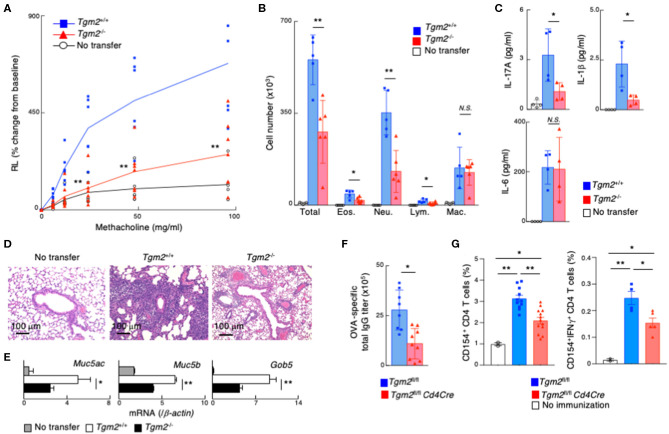
Memory helper T (Th) cells are crucial for secondary immune responses against infectious microorganisms but also drive the pathogenesis of chronic inflammatory diseases. Therefore, it is of fundamental importance to understand how memory T cells are generated. However, the molecular mechanisms governing memory Th cell generation remain incompletely understood. Here, we identified CD30 as a molecule heterogeneously expressed on effector Th1 and Th17 cells, and CD30hi effector Th1 and Th17 cells preferentially generated memory Th1 and Th17 cells. We found that CD30 mediated signal induced Transglutaminase-2 (TG2) expression, and that the TG2 expression in effector Th cells is essential for memory Th cell generation. In fact, Cd30-deficiency resulted in the impaired generation of memory Th1 and Th17 cells, which can be rescued by overexpression of TG2. Furthermore, transglutaminase-2 (Tgm2)-deficient CD4 T cells failed to become memory Th cells. As a result, T cells from Tgm2-deficient mice displayed impaired antigen-specific antibody production and attenuated Th17-mediated allergic responses. Our data indicate that CD30-induced TG2 expression in effector Th cells is essential for the generation of memory Th1 and Th17 cells, and that CD30 can be a marker for precursors of memory Th1 and Th17 cells.
Keywords: CD30; TG2; Th1; Th17; airway inflammation; memory Th cell generation; memory precursor.
Copyright © 2020 Suzuki, Yagi, Kimura, Iwamura, Shinoda, Onodera, Hirahara, Tumes, Koyama-Nasu, Iismaa, Graham, Motohashi and Nakayama.
Figures
Similar articles
-
Inositol polyphosphate multikinase regulates Th1 and Th17 cell differentiation by controlling Akt-mTOR signaling.Cell Rep. 2025 Feb 25;44(2):115281. doi: 10.1016/j.celrep.2025.115281. Epub 2025 Feb 12. Cell Rep. 2025. PMID: 39946233
-
CD30 ligand is a new therapeutic target for central nervous system autoimmunity.J Autoimmun. 2015 Feb;57:14-23. doi: 10.1016/j.jaut.2014.11.005. Epub 2014 Dec 19. J Autoimmun. 2015. PMID: 25533628
-
IL-21 is a central memory T cell-associated cytokine that inhibits the generation of pathogenic Th1/17 effector cells.J Immunol. 2014 Oct 1;193(7):3322-31. doi: 10.4049/jimmunol.1400775. Epub 2014 Aug 29. J Immunol. 2014. PMID: 25172491 Clinical Trial.
-
Origins of CD4(+) effector and central memory T cells.Nat Immunol. 2011 Jun;12(6):467-71. doi: 10.1038/ni.2038. Nat Immunol. 2011. PMID: 21739668 Free PMC article. Review.
-
The Multifaceted Role of Th1, Th9, and Th17 Cells in Immune Checkpoint Inhibition Therapy.Front Immunol. 2021 Mar 12;12:625667. doi: 10.3389/fimmu.2021.625667. eCollection 2021. Front Immunol. 2021. PMID: 33777008 Free PMC article. Review.
Cited by
-
Comparison of CD30L and OX40L Reveals CD30L as a Promising Therapeutic Target in Atopic Dermatitis.Allergy. 2025 Feb;80(2):500-512. doi: 10.1111/all.16412. Epub 2024 Nov 26. Allergy. 2025. PMID: 39589186 Free PMC article.
-
Advances and challenges in identifying precursors of memory CD4+ T cells.Front Immunol. 2025 May 5;16:1540045. doi: 10.3389/fimmu.2025.1540045. eCollection 2025. Front Immunol. 2025. PMID: 40391228 Free PMC article. Review.
-
Recent Thymic Emigrants Require RBPJ-Dependent Notch Signaling to Transition into Functionally Mature Naive T Cells.J Immunol. 2023 Jul 1;211(1):81-90. doi: 10.4049/jimmunol.2300140. J Immunol. 2023. PMID: 37154711 Free PMC article.
-
Role of Transglutaminase 2 in Cell Death, Survival, and Fibrosis.Cells. 2021 Jul 20;10(7):1842. doi: 10.3390/cells10071842. Cells. 2021. PMID: 34360011 Free PMC article. Review.
-
The Role of Transglutaminase 2 in Cancer: An Update.Int J Mol Sci. 2024 Feb 28;25(5):2797. doi: 10.3390/ijms25052797. Int J Mol Sci. 2024. PMID: 38474044 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous