

The Impact of Malaria Parasites on Dendritic Cell-T Cell Interaction
- PMID: 32793231
- PMCID: PMC7393936
- DOI: 10.3389/fimmu.2020.01597
The Impact of Malaria Parasites on Dendritic Cell-T Cell Interaction
Abstract
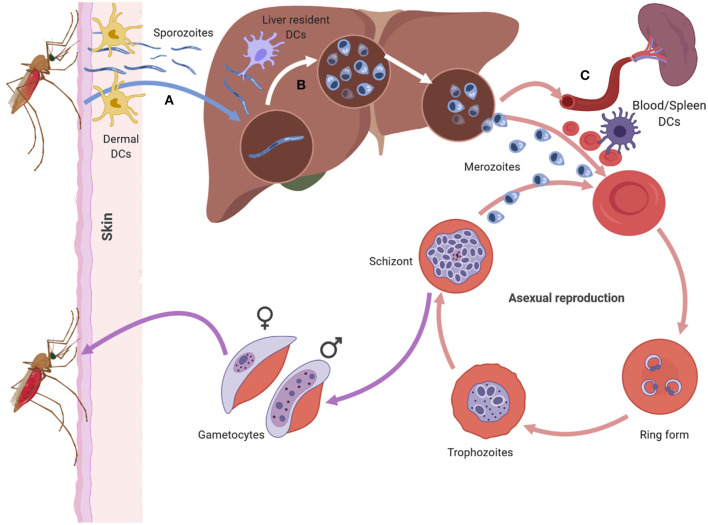
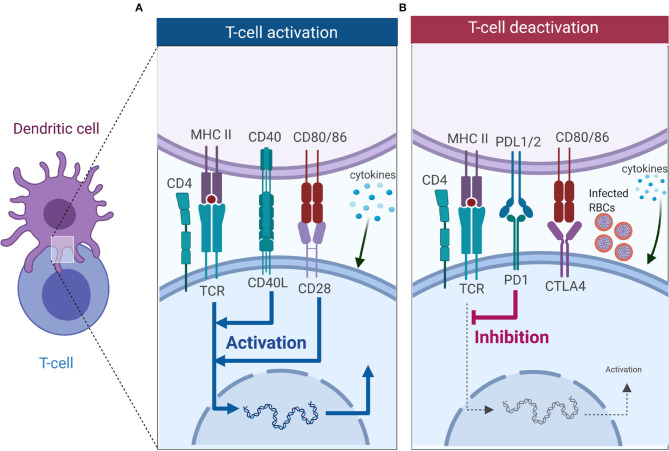
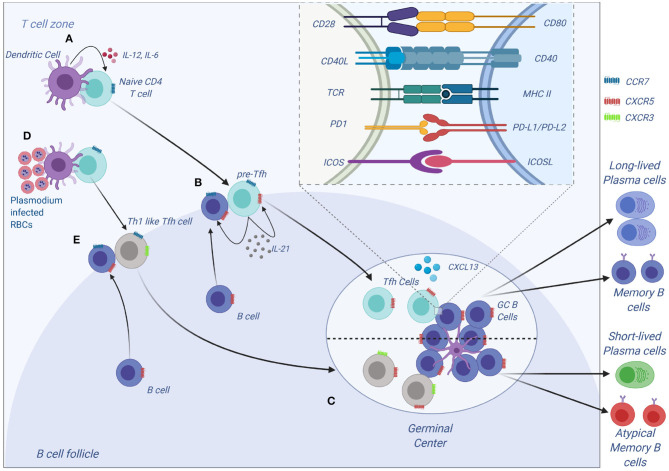
Malaria is caused by apicomplexan parasites of the genus Plasmodium. While infection continues to pose a risk for the majority of the global population, the burden of disease mainly resides in Sub-Saharan Africa. Although immunity develops against disease, this requires years of persistent exposure and is not associated with protection against infection. Repeat infections occur due to the parasite's ability to disrupt or evade the host immune responses. However, despite many years of study, the mechanisms of this disruption remain unclear. Previous studies have demonstrated a parasite-induced failure in dendritic cell (DCs) function affecting the generation of helper T cell responses. These T cells fail to help B cell responses, reducing the production of antibodies that are necessary to control malaria infection. This review focuses on our current understanding of the effect of Plasmodium parasite on DC function, DC-T cell interaction, and T cell activation. A better understanding of how parasites disrupt DC-T cell interactions will lead to new targets and approaches to reinstate adaptive immune responses and enhance parasite immunity.
Keywords: DC-T cell interaction; T cells; T helper cells; Tfh; dendritic cells; malaria.
Copyright © 2020 Osii, Otto, Garside, Ndungu and Brewer.
Figures
References
-
- WHO World Malaria Report 2018 (2018).
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical