

HEARTBREAK Controls Post-translational Modification of INDEHISCENT to Regulate Fruit Morphology in Capsella
- PMID: 32795439
- PMCID: PMC7544509
- DOI: 10.1016/j.cub.2020.07.055
HEARTBREAK Controls Post-translational Modification of INDEHISCENT to Regulate Fruit Morphology in Capsella
Abstract
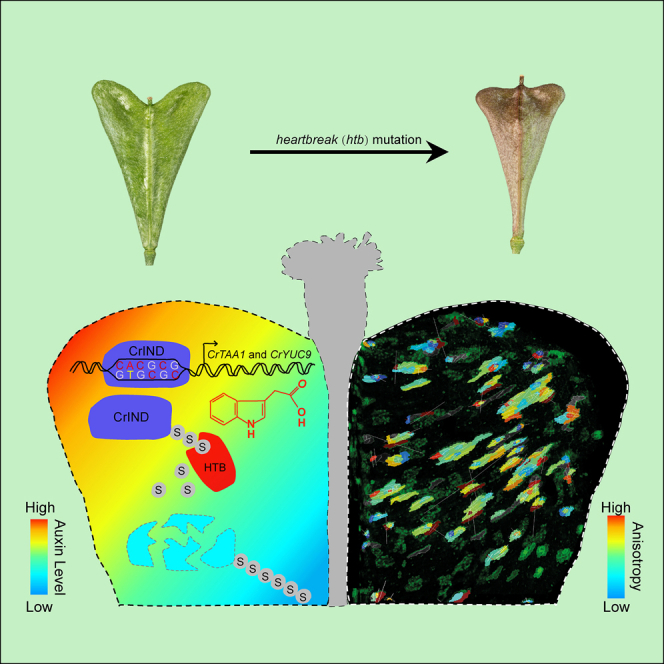
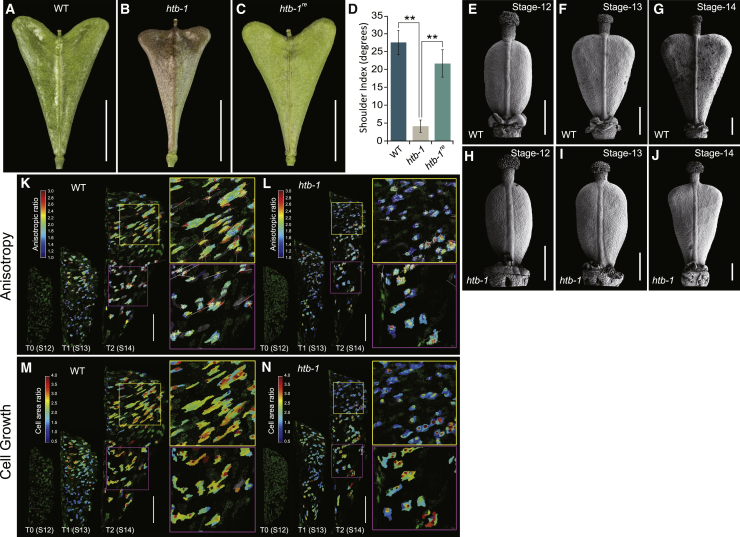
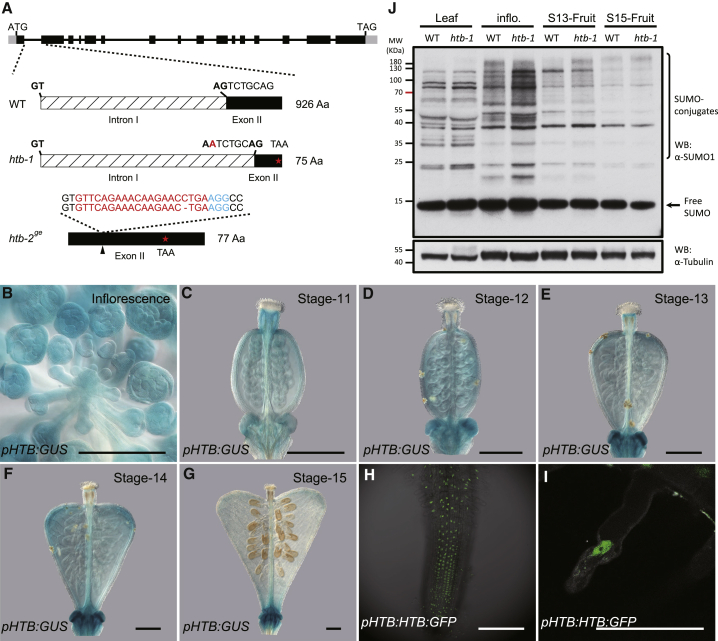
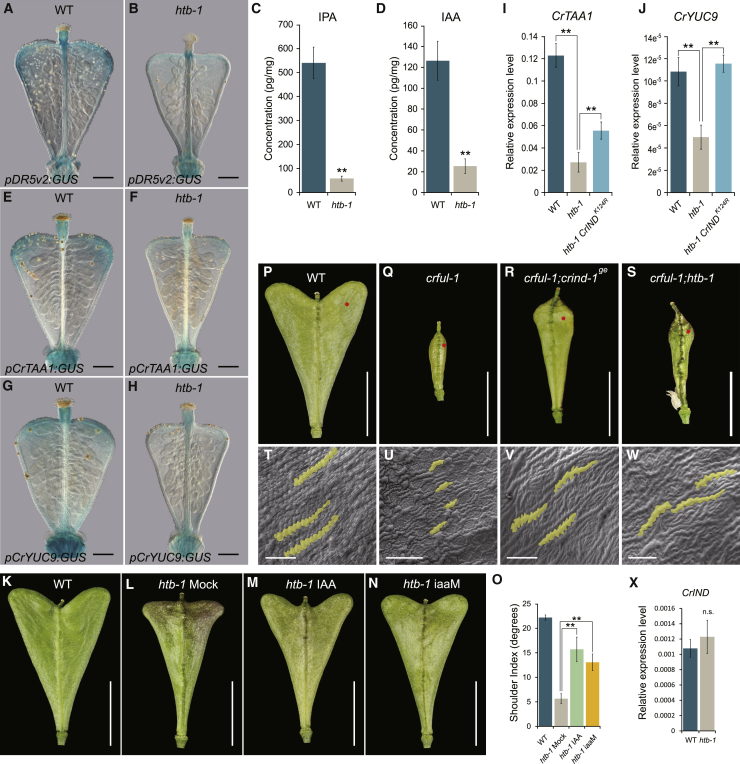
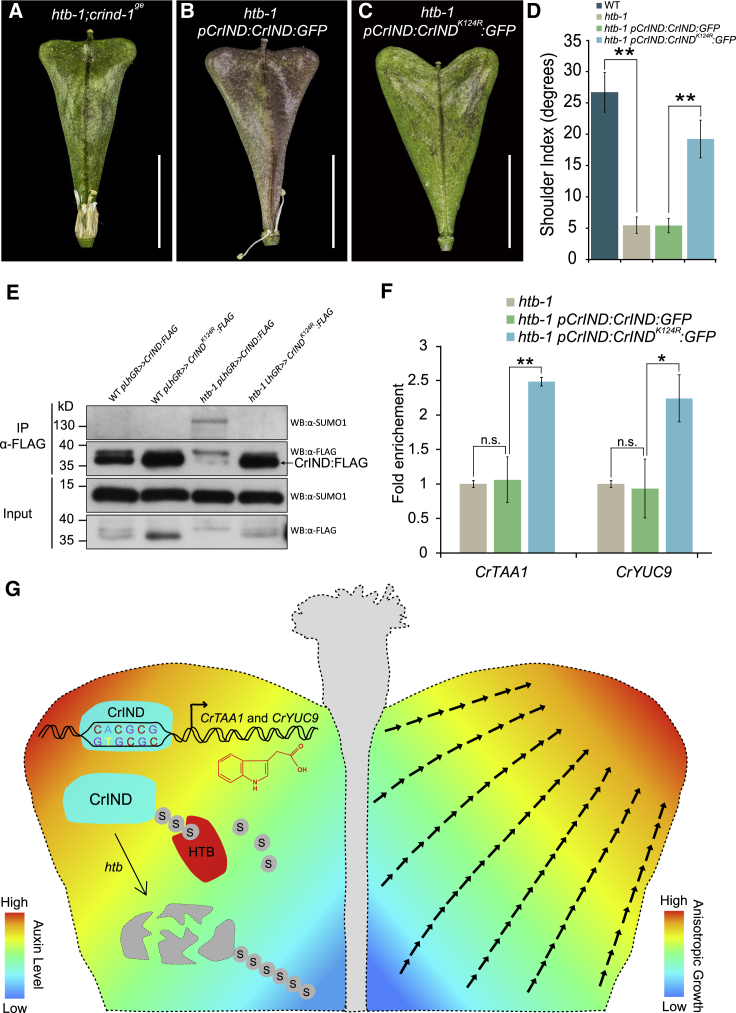
Morphological variation is the basis of natural diversity and adaptation. For example, angiosperms (flowering plants) evolved during the Cretaceous period more than 100 mya and quickly colonized terrestrial habitats [1]. A major reason for their astonishing success was the formation of fruits, which exist in a myriad of different shapes and sizes [2]. Evolution of organ shape is fueled by variation in expression patterns of regulatory genes causing changes in anisotropic cell expansion and division patterns [3-5]. However, the molecular mechanisms that alter the polarity of growth to generate novel shapes are largely unknown. The heart-shaped fruits produced by members of the Capsella genus comprise an anatomical novelty, making it particularly well suited for studies on morphological diversification [6-8]. Here, we show that post-translational modification of regulatory proteins provides a critical step in organ-shape formation. Our data reveal that the SUMO protease, HEARTBREAK (HTB), from Capsella rubella controls the activity of the key regulator of fruit development, INDEHISCENT (CrIND in C. rubella), via de-SUMOylation. This post-translational modification initiates a transduction pathway required to ensure precisely localized auxin biosynthesis, thereby facilitating anisotropic cell expansion to ultimately form the heart-shaped Capsella fruit. Therefore, although variation in the expression of key regulatory genes is known to be a primary driver in morphological evolution, our work demonstrates how other processes-such as post-translational modification of one such regulator-affects organ morphology.
Keywords: Capsella rubella; SUMOylation; anisotropic cell growth; fruit morphology; gene expression; post-translational modification.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Soltis P.S., Folk R.A., Soltis D.E. Darwin review: angiosperm phylogeny and evolutionary radiations. Proc. Royal Soc. B: Biol. Sci. 2019;286:20190099.
-
- Seymour G.B., Østergaard L., Chapman N.H., Knapp S., Martin C. Fruit development and ripening. Annu. Rev. Plant Biol. 2013;64:219–241. - PubMed
-
- Carroll S.B. Endless forms: the evolution of gene regulation and morphological diversity. Cell. 2000;101:577–580. - PubMed
-
- Wray G.A. The evolutionary significance of cis-regulatory mutations. Nat. Rev. Genet. 2007;8:206–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
