

Interrogating Cellular Communication in Cancer with Genetically Encoded Imaging Reporters
- PMID: 32803164
- PMCID: PMC7398120
- DOI: 10.1148/rycan.2020190053
Interrogating Cellular Communication in Cancer with Genetically Encoded Imaging Reporters
Abstract
Cells continuously communicate changes in their microenvironment, both locally and globally, with other cells in the organism. Integration of information arising from signaling networks impart continuous, time-dependent changes of cell function and phenotype. Use of genetically encoded reporters enable researchers to noninvasively monitor time-dependent changes in intercellular and intracellular signaling, which can be interrogated by macroscopic and microscopic optical imaging, nuclear medicine imaging, MRI, and even photoacoustic imaging techniques. Reporters enable noninvasive monitoring of changes in cell-to-cell proximity, transcription, translation, protein folding, protein association, protein degradation, drug action, and second messengers in real time. Because of their positive impact on preclinical research, attempts to improve the sensitivity and specificity of these reporters, and to develop new types and classes of reporters, remain an active area of investigation. A few reporters have migrated to proof-of-principle clinical demonstrations, and recent advances in genome editing technologies may enable the use of reporters in the context of genome-wide analysis and the imaging of complex genomic regulation in vivo that cannot be readily investigated through standard methodologies. The combination of genetically encoded imaging reporters with continuous improvements in other molecular biology techniques may enhance and expedite target discovery and drug development for cancer interventions and treatment. © RSNA, 2020.
2020 by the Radiological Society of North America, Inc.
Figures
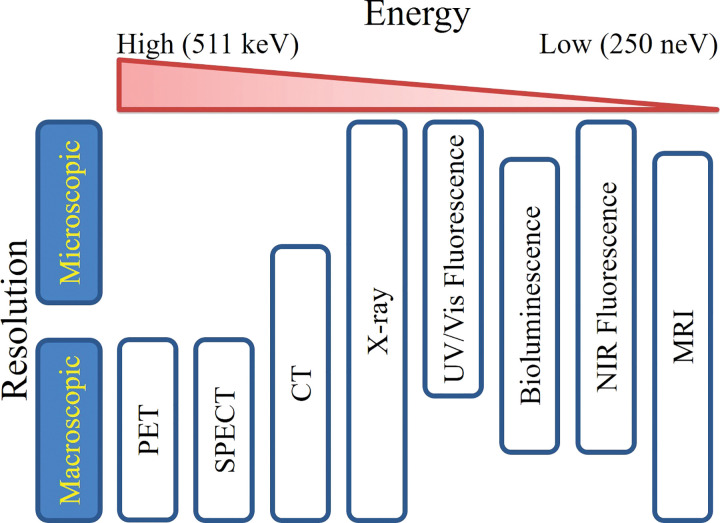
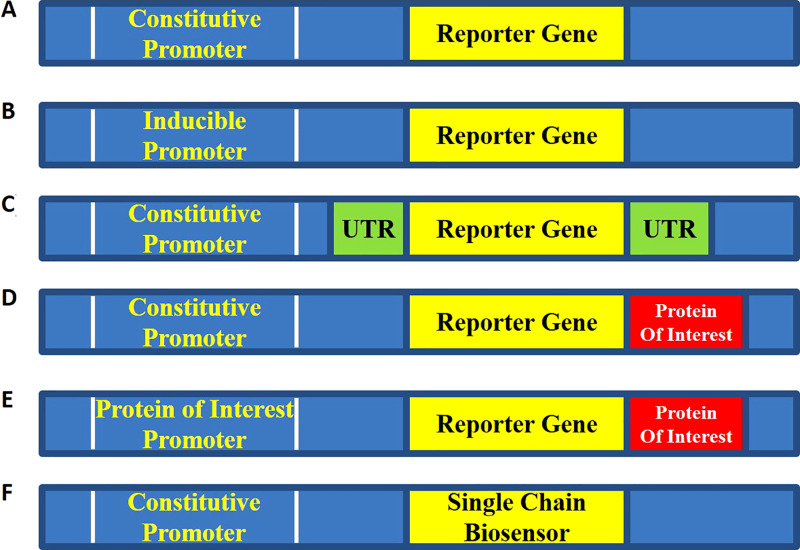
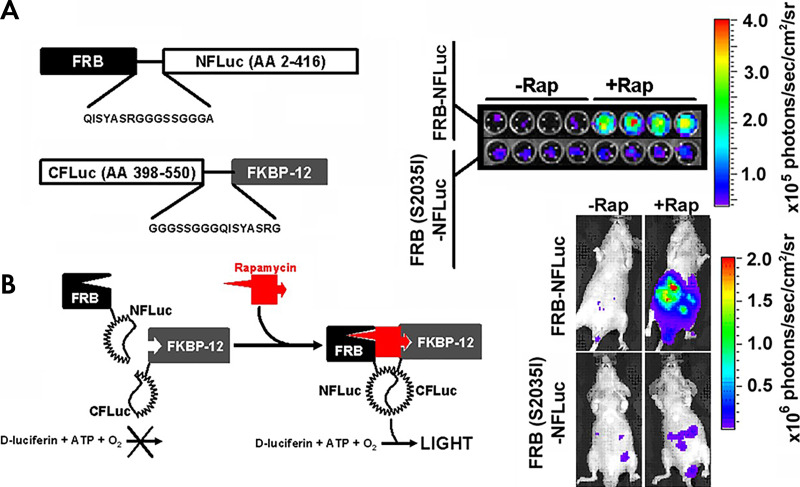
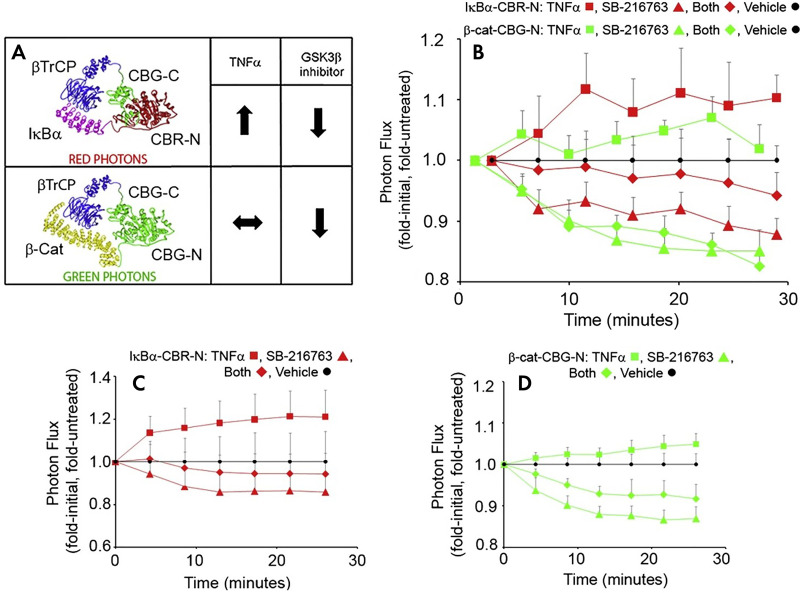
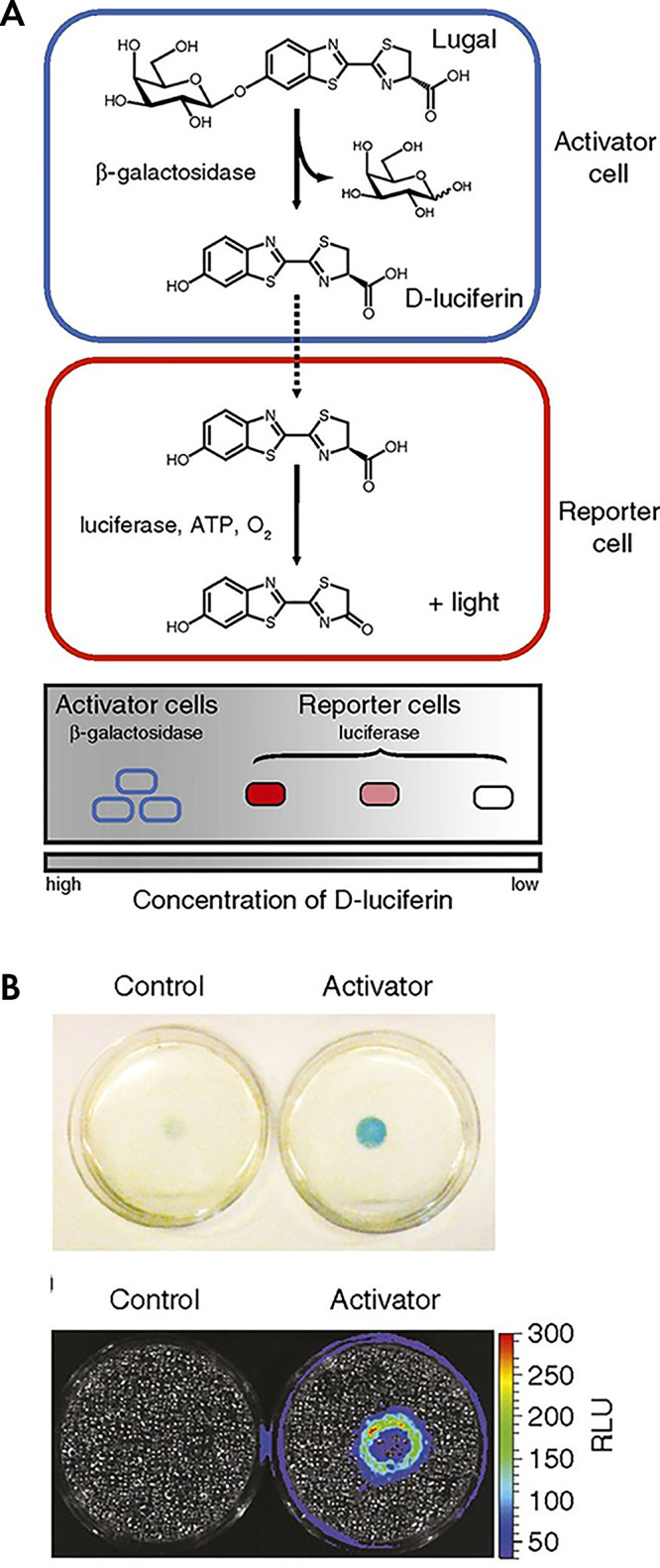
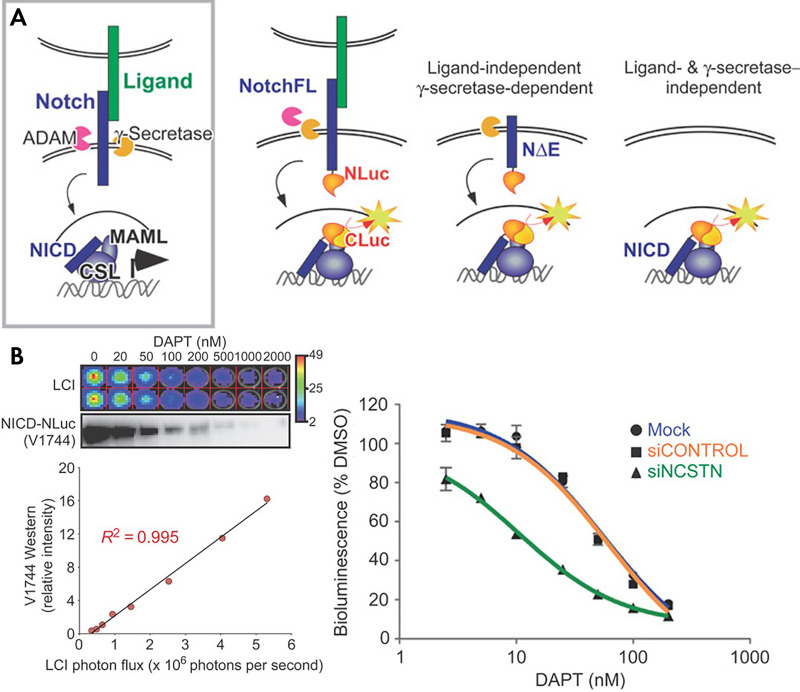
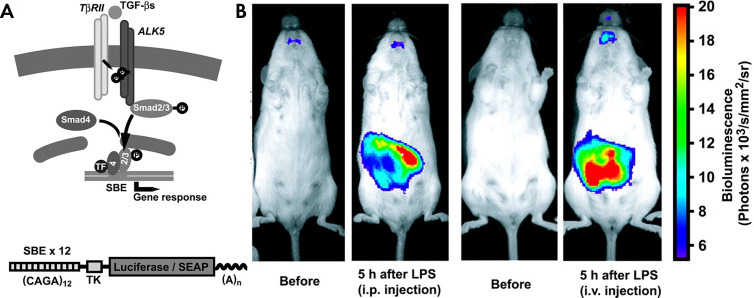
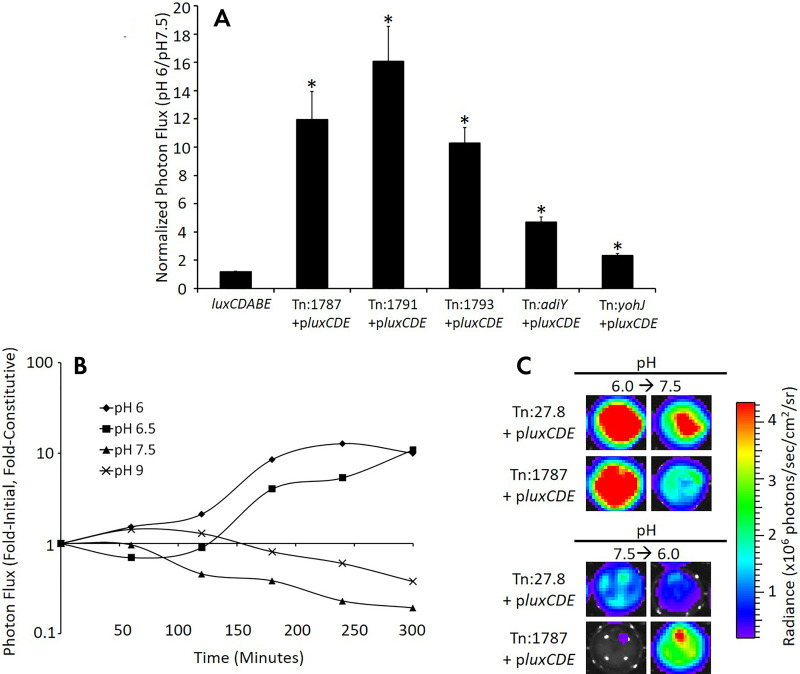
References
-
- O’Connor CM, Adams JU. Essentials of Cell Biology. Cambridge, Mass: NPG Education, 2010.
-
- Couzin-Frankel J. Breakthrough of the year 2013. Cancer immunotherapy. Science 2013;342(6165):1432–1433. - PubMed
-
- Wei SC, Duffy CR, Allison JP. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov 2018;8(9):1069–1086. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
