

Mitochondrial fatty acid synthesis coordinates oxidative metabolism in mammalian mitochondria
- PMID: 32804083
- PMCID: PMC7470841
- DOI: 10.7554/eLife.58041
Mitochondrial fatty acid synthesis coordinates oxidative metabolism in mammalian mitochondria
Abstract
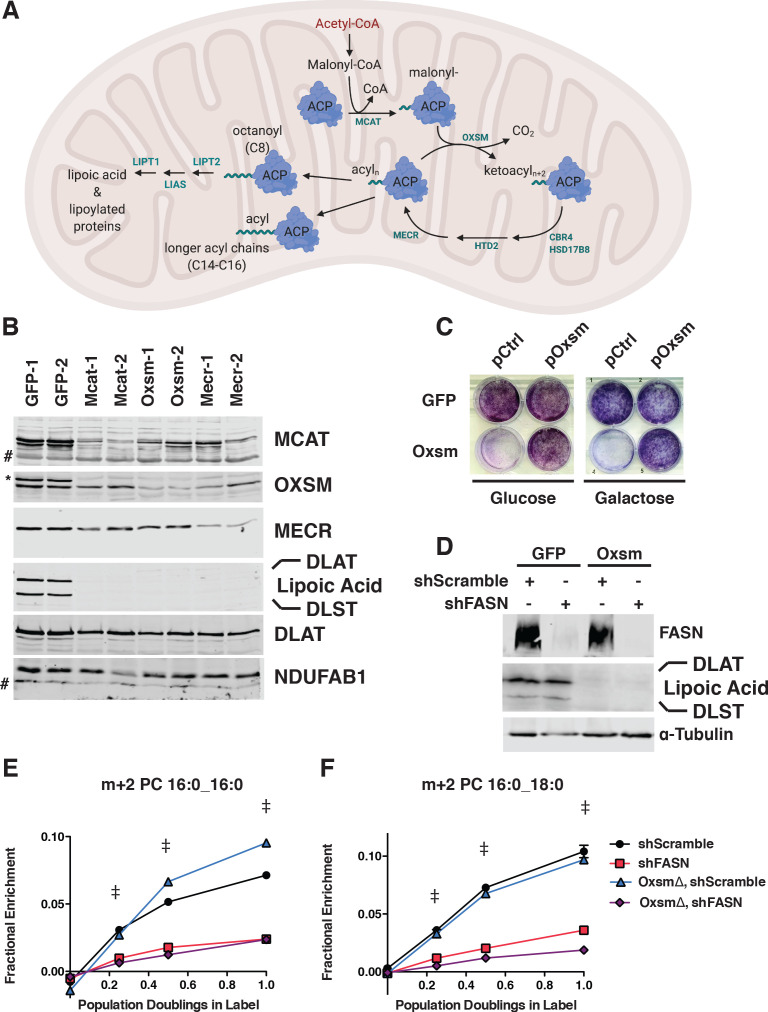
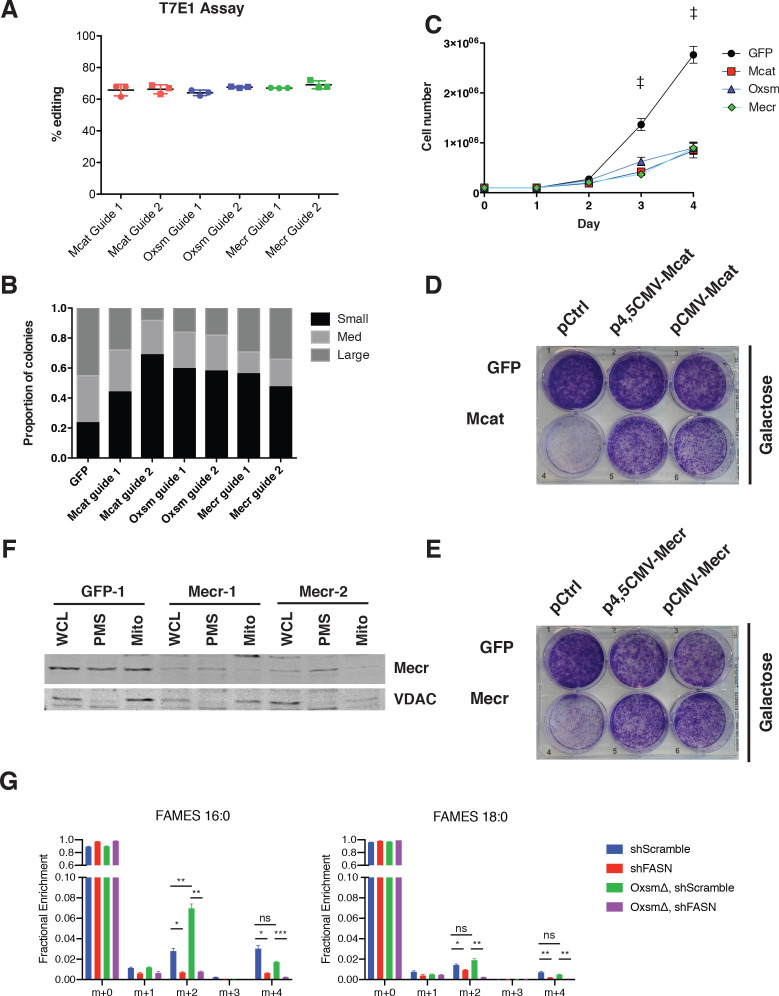
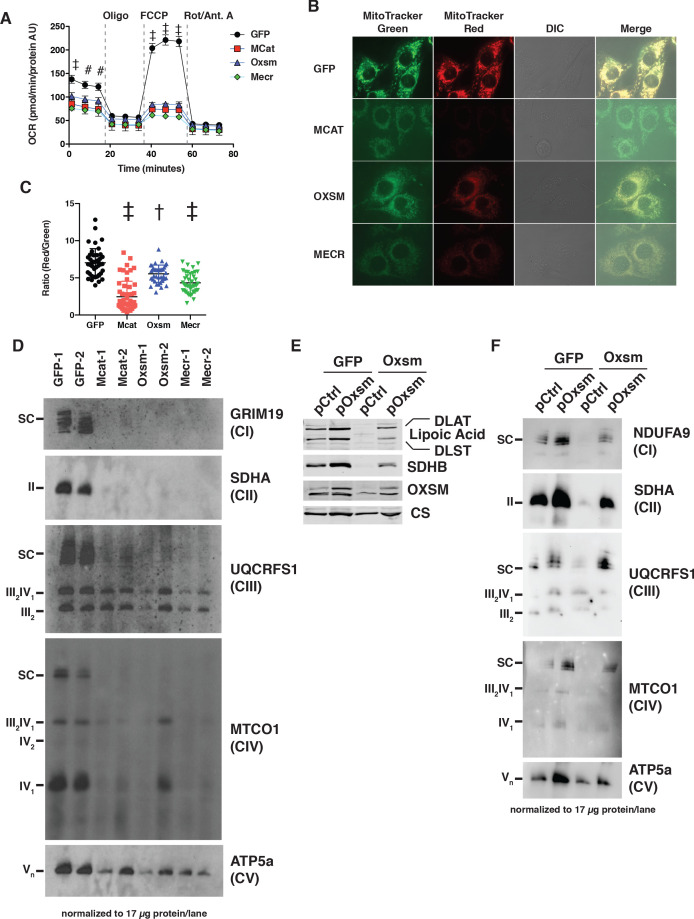
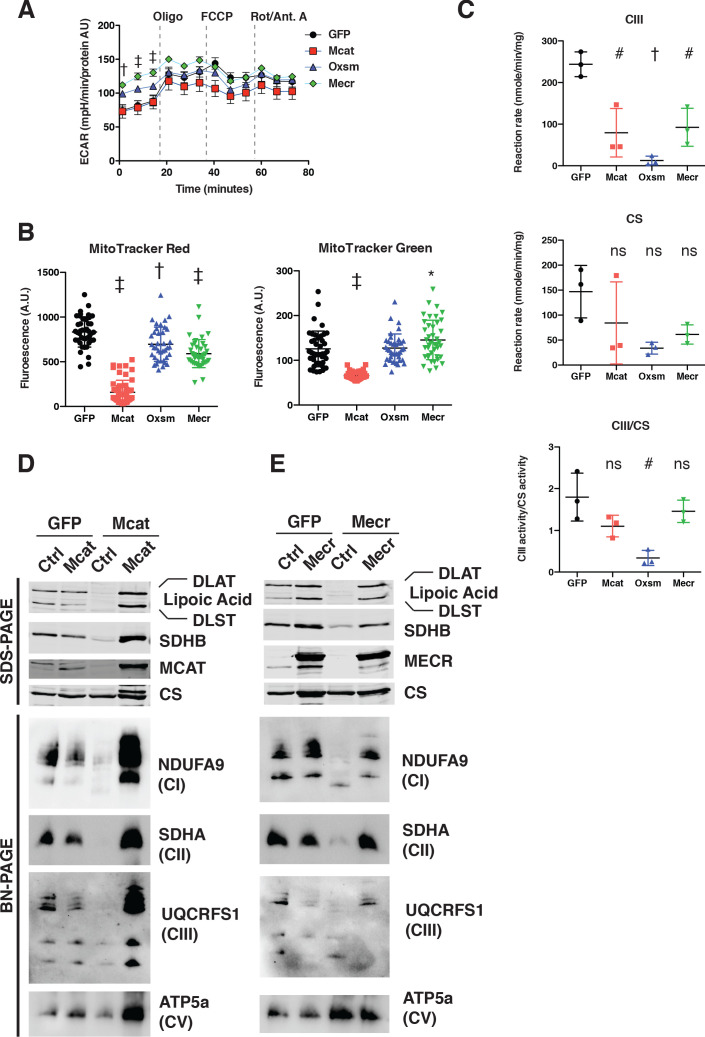
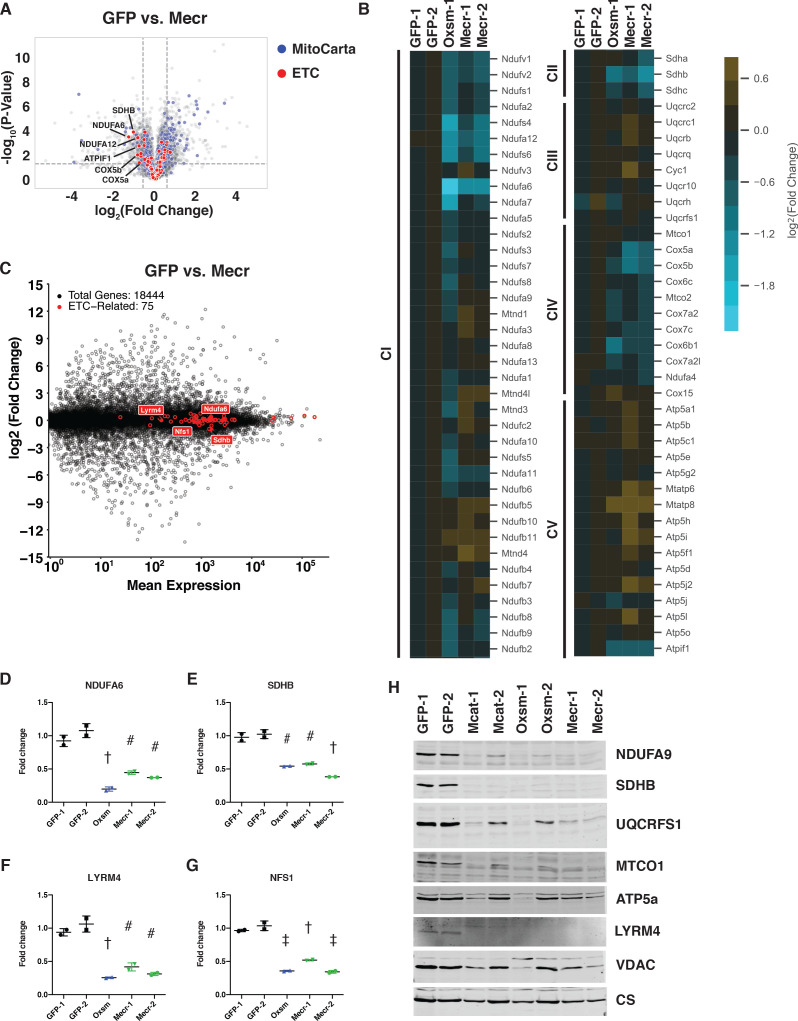
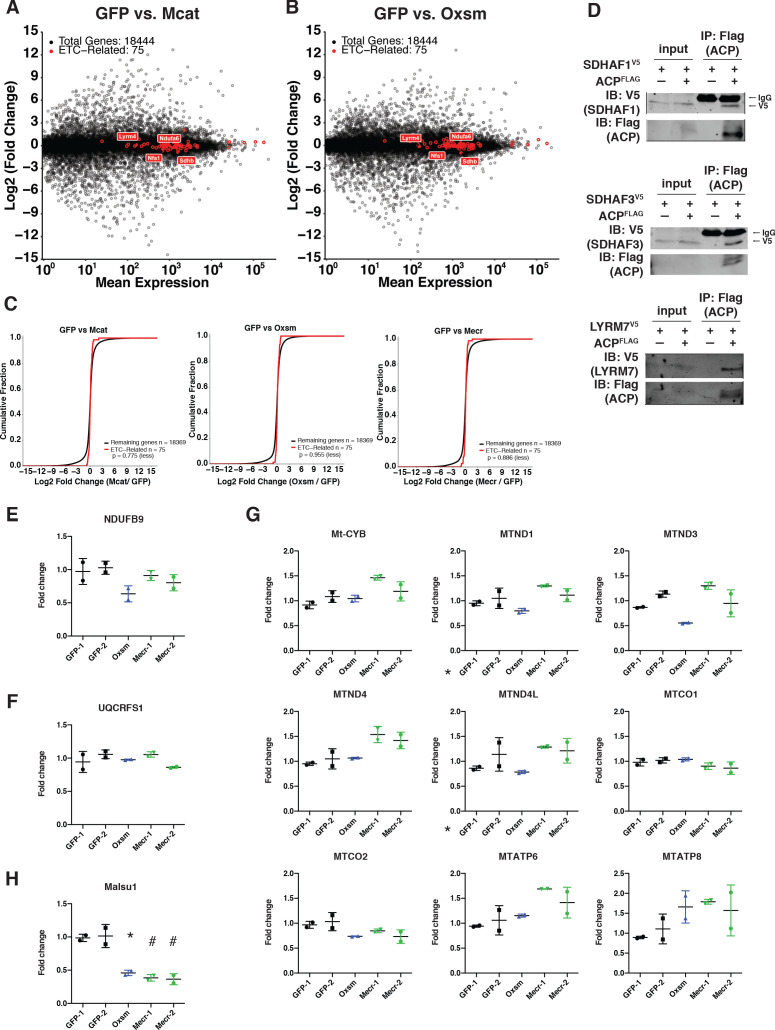
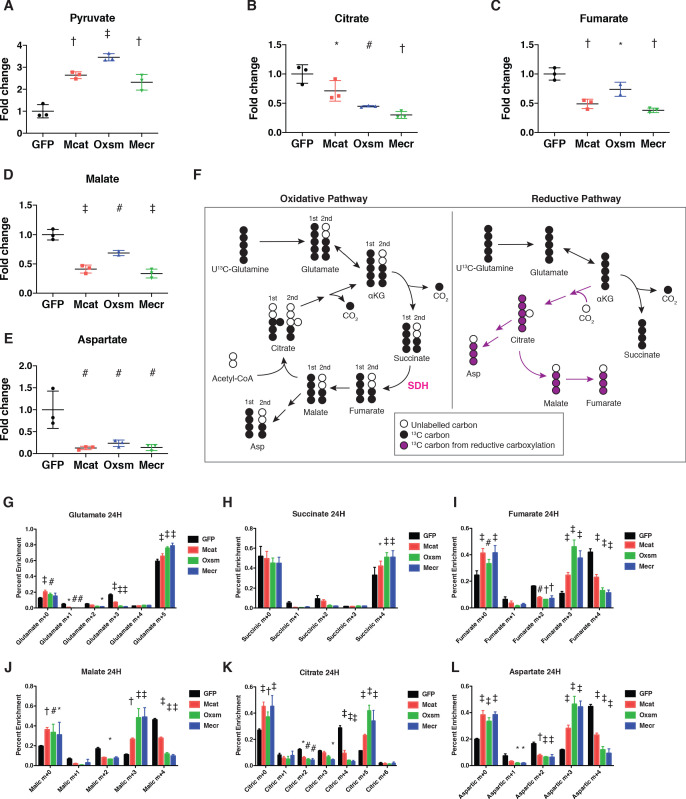
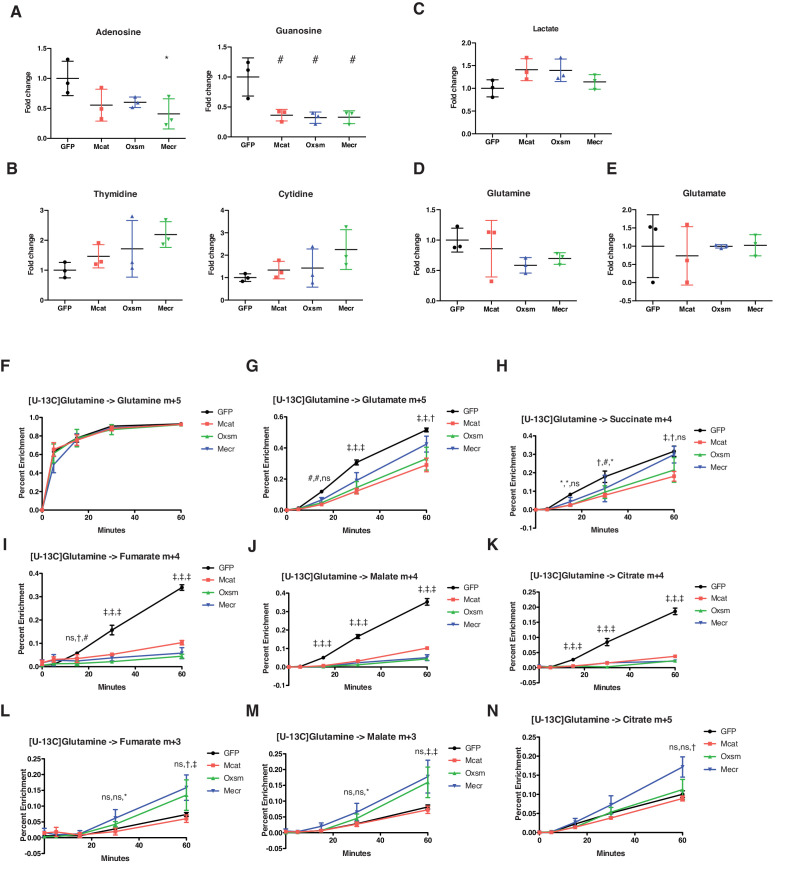
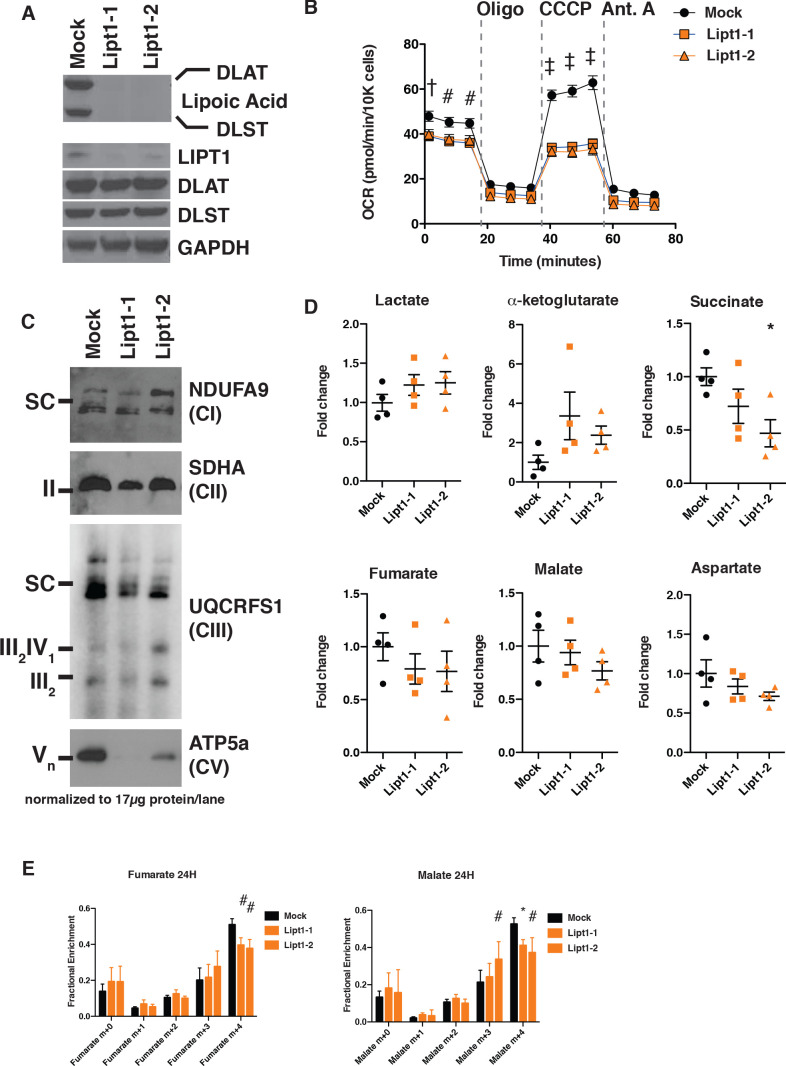
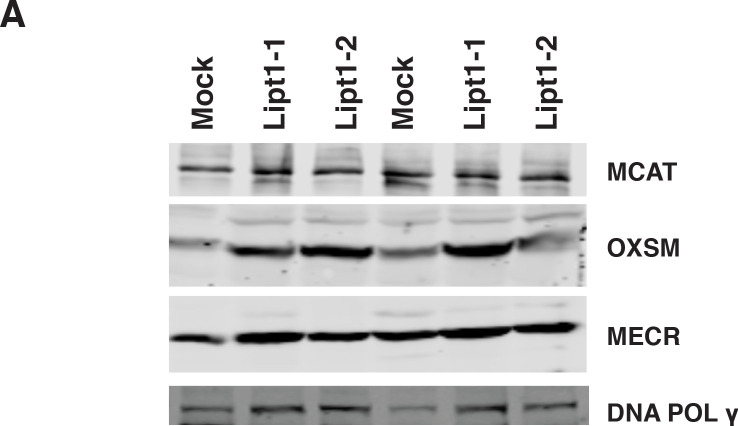
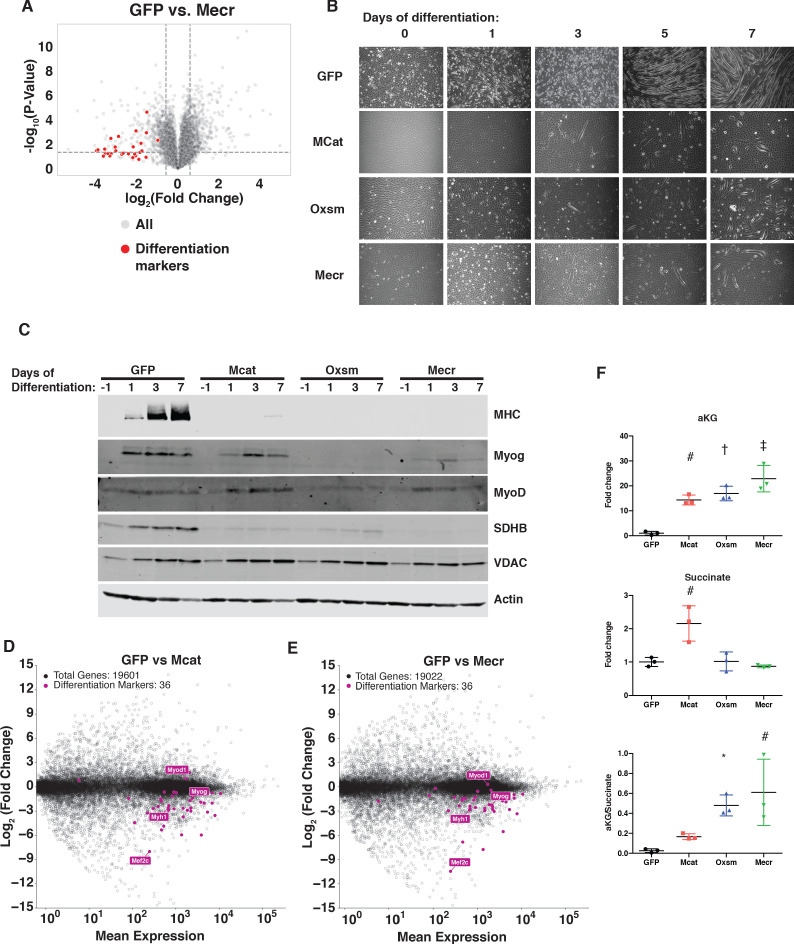
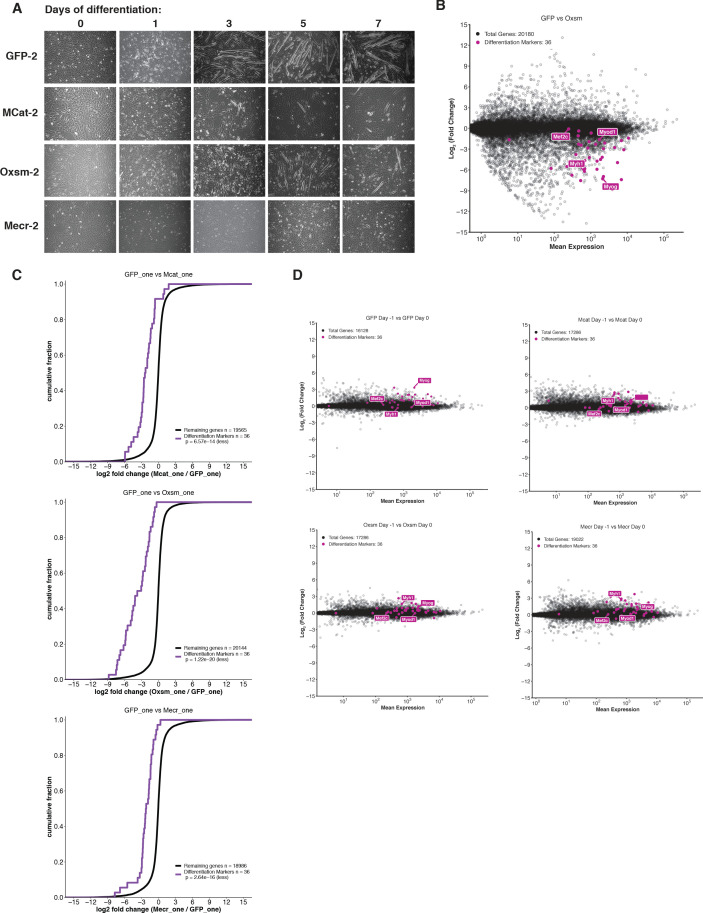
Cells harbor two systems for fatty acid synthesis, one in the cytoplasm (catalyzed by fatty acid synthase, FASN) and one in the mitochondria (mtFAS). In contrast to FASN, mtFAS is poorly characterized, especially in higher eukaryotes, with the major product(s), metabolic roles, and cellular function(s) being essentially unknown. Here we show that hypomorphic mtFAS mutant mouse skeletal myoblast cell lines display a severe loss of electron transport chain (ETC) complexes and exhibit compensatory metabolic activities including reductive carboxylation. This effect on ETC complexes appears to be independent of protein lipoylation, the best characterized function of mtFAS, as mutants lacking lipoylation have an intact ETC. Finally, mtFAS impairment blocks the differentiation of skeletal myoblasts in vitro. Together, these data suggest that ETC activity in mammals is profoundly controlled by mtFAS function, thereby connecting anabolic fatty acid synthesis with the oxidation of carbon fuels.
Keywords: TCA cycle; biochemistry; chemical biology; electron transport chain; fatty acid synthesis; metabolism; mitochondria; mouse; myoblast.
Plain language summary
In human, plant and other eukaryotic cells, fats are an important source of energy and also play many other roles including waterproofing, thermal insulation and energy storage. Eukaryotic cells have two systems that make the building blocks of fats (known as fatty acids) and one of these systems, called the mtFAS pathway, operates in small compartments known as mitochondria. This pathway only has one known product, a small fat molecule called lipoic acid, which mitochondria attach to several enzymes to allow them to work properly. The main role of mitochondria is to break down fats and other molecules to release chemical energy that powers many processes in cells. They achieve this using large groups of proteins known as ETC complexes. To build these complexes, families of proteins known as ETC assembly factors carefully coordinate the assembly of many proteins and small molecules into specific structures. However, it remains unclear precisely how this process works. Here, Nowinski et al. used a gene editing technique to mutate the genes encoding three enzymes in the mtFAS pathway in mammalian cells. The experiments found that the mutant cells had fewer ETC complexes and seemed to be less able to break down fats and other molecules than ‘normal’ cells. Furthermore, a family of ETC assembly factors were less stable in the mutant cells. These findings suggest that the mtFAS pathway controls how mitochondria assemble ETC complexes. Further experiments indicated that lipoic acid is not involved in the assembly of ETC complexes and that the mtFAS pathway produces another, as yet unidentified, product that regulates this process, instead. MEPAN syndrome is a rare neurological disorder that leads to progressive loss of control of movement, slurred speech and impaired vision in children. Patients with this syndrome have genetic mutations affecting components of the mtFAS pathway, therefore, a better understanding of how the pathway works may help researchers develop new treatments in the future. More broadly, these findings will have important ramifications for many other situations in which the activity of ETC complexes in mitochondria is modified.
© 2020, Nowinski et al.
Conflict of interest statement
SN, AS, SR, JM, CB, SF, MJ, SL, JB, JM, YO, BN, JP, KF, JC, SG, DW, JR No competing interests declared, RD Reviewing editor, eLife
Figures
References
-
- Angerer H, Radermacher M, Mańkowska M, Steger M, Zwicker K, Heide H, Wittig I, Brandt U, Zickermann V. The LYR protein subunit NB4M/NDUFA6 of mitochondrial complex I anchors an acyl carrier protein and is essential for catalytic activity. PNAS. 2014;111:5207–5212. doi: 10.1073/pnas.1322438111. - DOI - PMC - PubMed
-
- Angerer H, Schönborn S, Gorka J, Bahr U, Karas M, Wittig I, Heidler J, Hoffmann J, Morgner N, Zickermann V. Acyl modification and binding of mitochondrial ACP to multiprotein complexes. Biochimica Et Biophysica Acta (BBA) - Molecular Cell Research. 2017;1864:1913–1920. doi: 10.1016/j.bbamcr.2017.08.006. - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- GM97645/GM/NIGMS NIH HHS/United States
- S10 OD016232/OD/NIH HHS/United States
- R35 GM131854/GM/NIGMS NIH HHS/United States
- S10 OD018210/OD/NIH HHS/United States
- R01 GM115174/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 GM110755/GM/NIGMS NIH HHS/United States
- R35 CA220449/CA/NCI NIH HHS/United States
- R21 AG063077/AG/NIA NIH HHS/United States
- T32 HL007576/HL/NHLBI NIH HHS/United States
- R01 GM132129/GM/NIGMS NIH HHS/United States
- R35CA22044901/CA/NCI NIH HHS/United States
- R01 DK107397/DK/NIDDK NIH HHS/United States
- U54 DK110858/DK/NIDDK NIH HHS/United States
- F32 HD096786/HD/NICHD NIH HHS/United States
- R01 GM067945/GM/NIGMS NIH HHS/United States
- T32DK11096601/DK/NIDDK NIH HHS/United States
- S10 OD021505/OD/NIH HHS/United States
- R01 GM115129/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
