

PDCD4 limits prooncogenic neuregulin-ErbB signaling
- PMID: 32804243
- PMCID: PMC11073242
- DOI: 10.1007/s00018-020-03617-5
PDCD4 limits prooncogenic neuregulin-ErbB signaling
Abstract
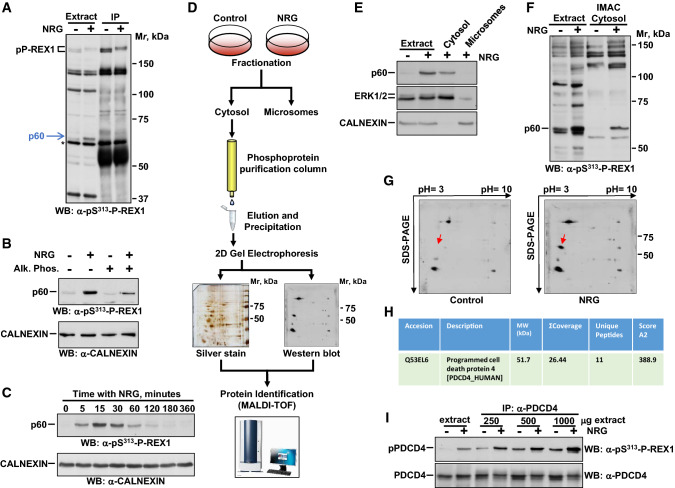
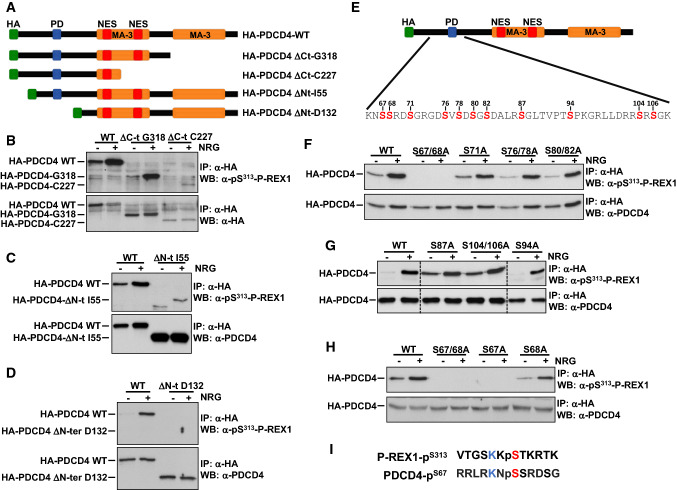
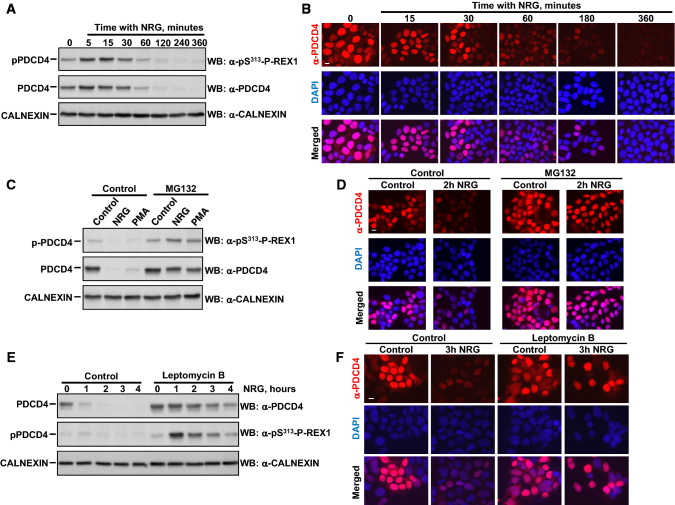
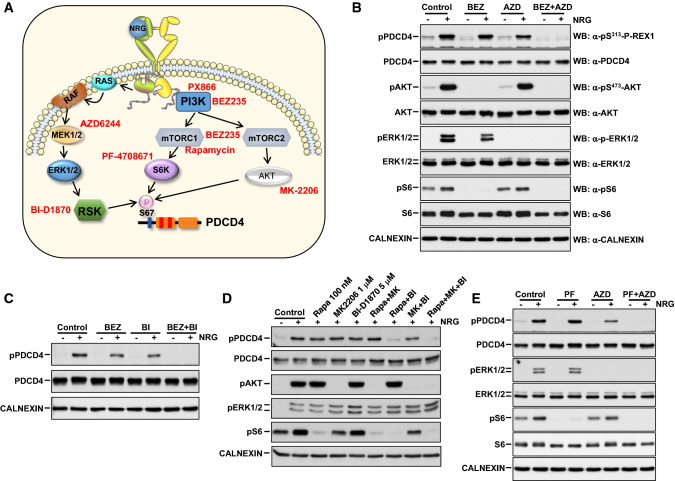
The neuregulins and their ErbB/HER receptors play essential roles in mammalian development and tissue homeostasis. In addition, deregulation of their function has been linked to the pathogenesis of diseases such as cancer or schizophrenia. These circumstances have stimulated research into the biology of this ligand-receptor system. Here we show the identification of programmed cell death protein-4 (PDCD4) as a novel neuregulin-ErbB signaling mediator. Phosphoproteomic analyses identified PDCD4 as protein whose phosphorylation increased in cells treated with neuregulin. Mutagenesis experiments defined serine 67 of PDCD4 as a site whose phosphorylation increased upon activation of neuregulin receptors. Phosphorylation of that site promoted degradation of PDCD4 by the proteasome, which depended on exit of PDCD4 from the nucleus to the cytosol. Mechanistic studies defined mTORC1 and ERK1/2 as routes implicated in neuregulin-induced serine 67 phosphorylation and PDCD4 degradation. Functionally, PDCD4 regulated several important biological functions of neuregulin, such as proliferation, migration, or invasion.
Keywords: Breast cancer; ErbB receptors; Neuregulin; PDCD4.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

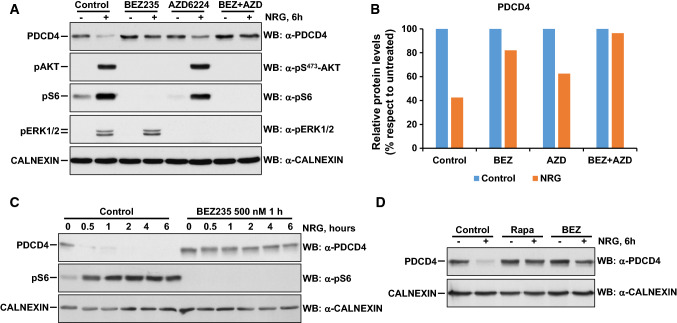
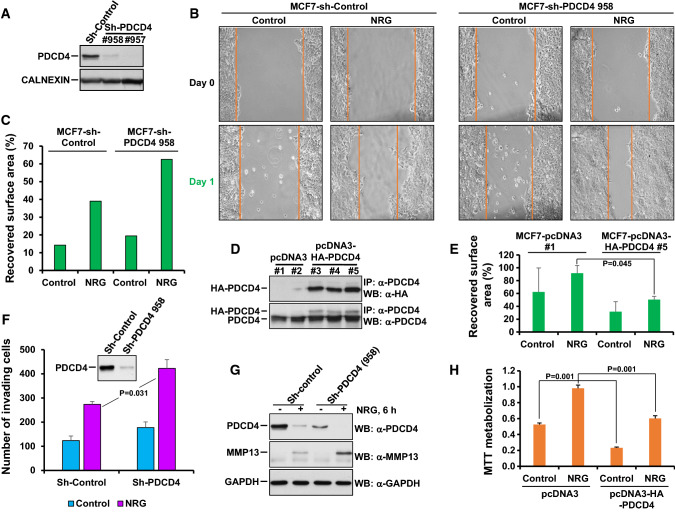
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
