

CDK12 globally stimulates RNA polymerase II transcription elongation and carboxyl-terminal domain phosphorylation
- PMID: 32805052
- PMCID: PMC7641311
- DOI: 10.1093/nar/gkaa514
CDK12 globally stimulates RNA polymerase II transcription elongation and carboxyl-terminal domain phosphorylation
Abstract
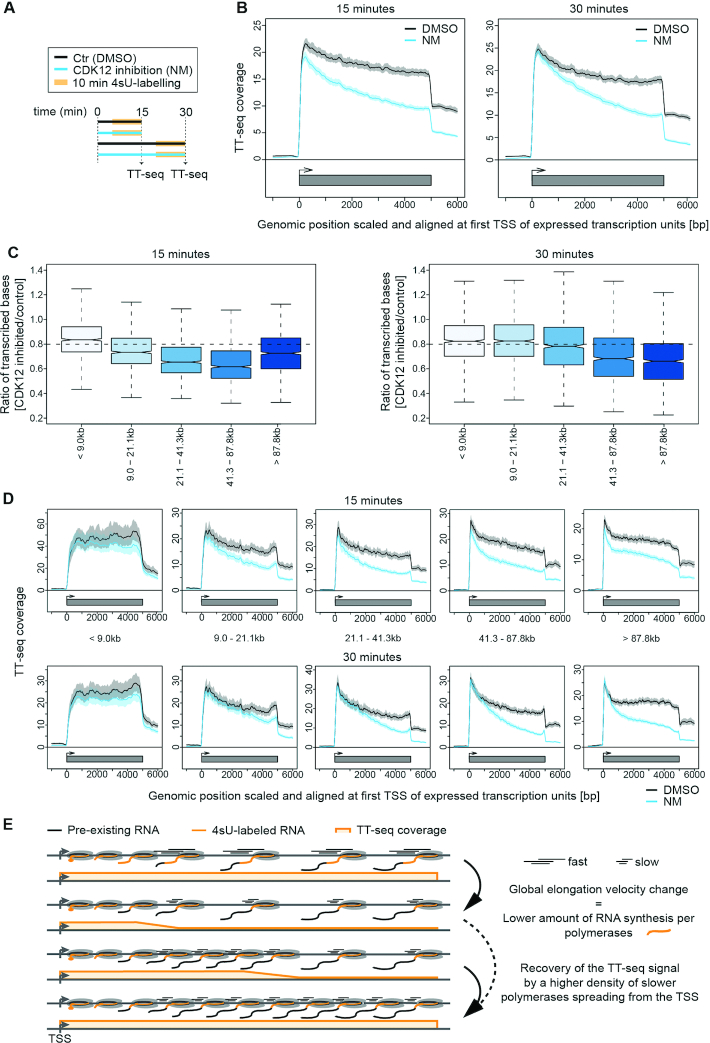
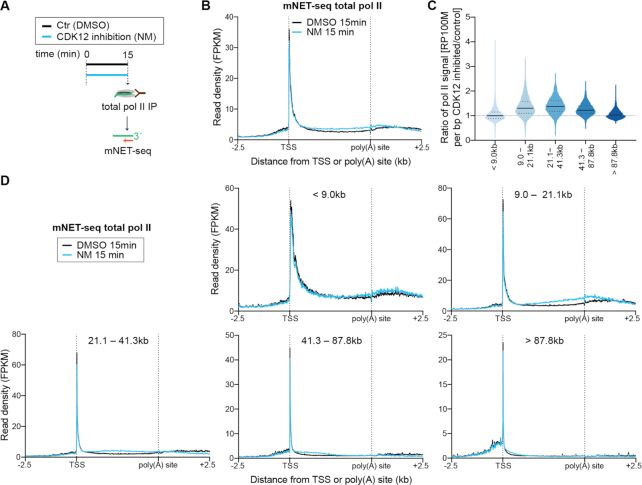
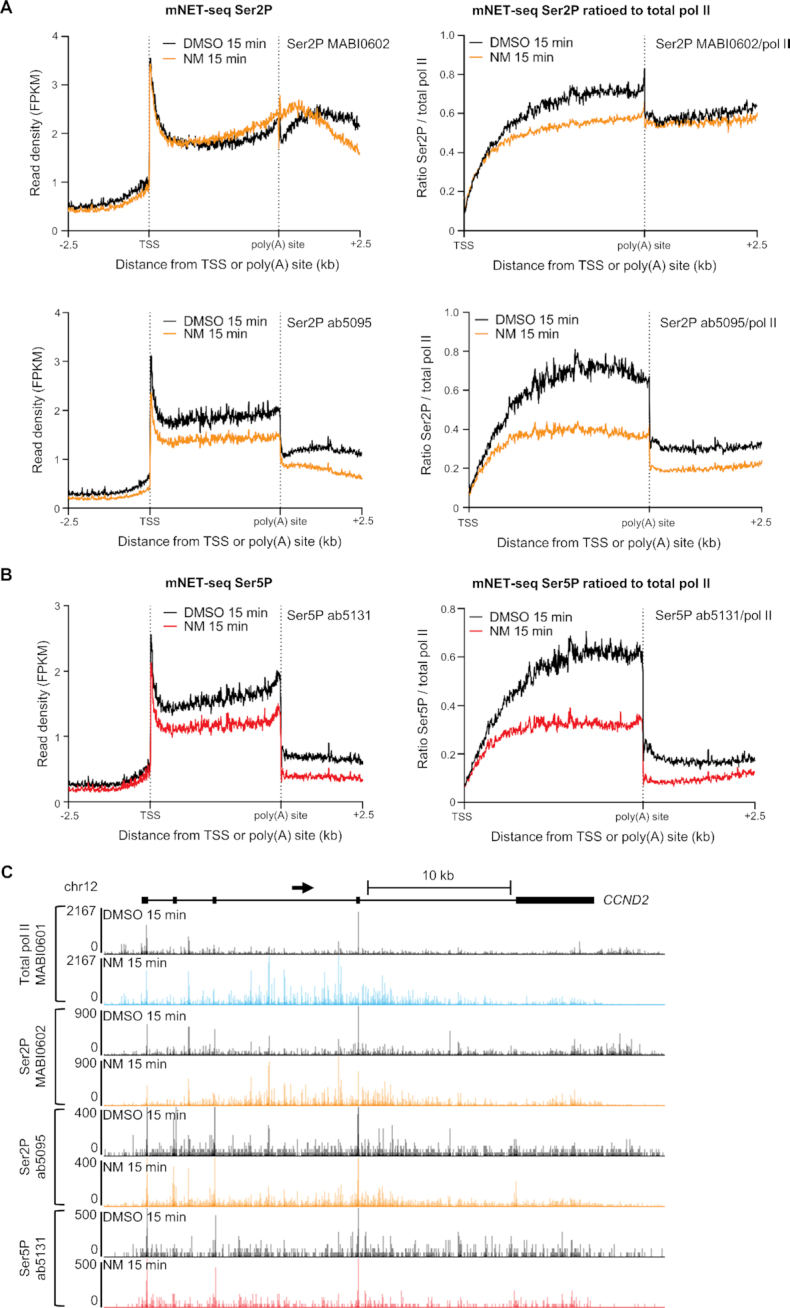
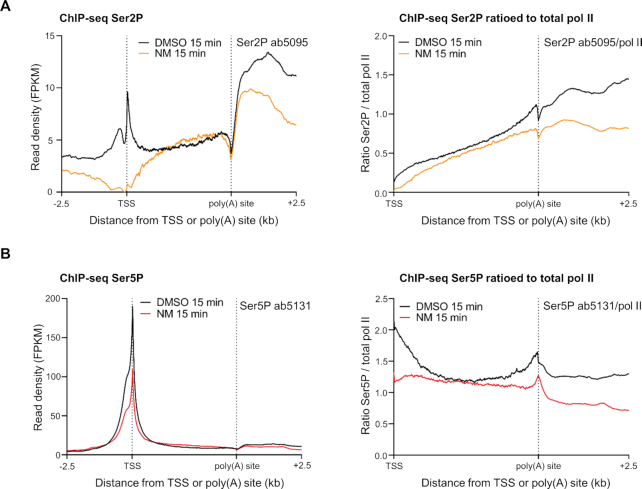
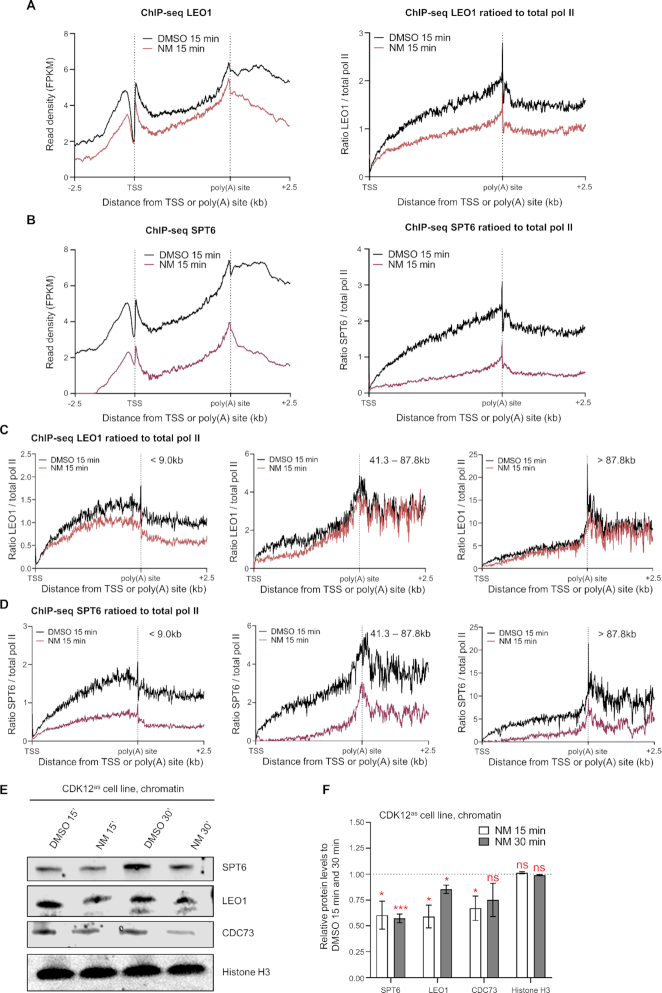
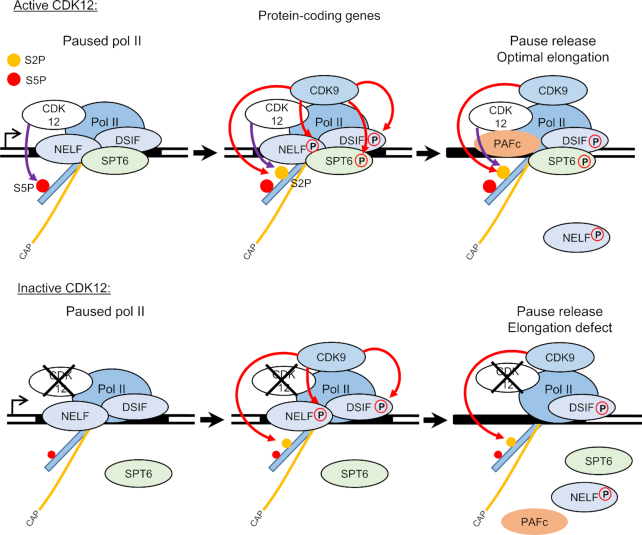
Cyclin-dependent kinase 12 (CDK12) phosphorylates the carboxyl-terminal domain (CTD) of RNA polymerase II (pol II) but its roles in transcription beyond the expression of DNA damage response genes remain unclear. Here, we have used TT-seq and mNET-seq to monitor the direct effects of rapid CDK12 inhibition on transcription activity and CTD phosphorylation in human cells. CDK12 inhibition causes a genome-wide defect in transcription elongation and a global reduction of CTD Ser2 and Ser5 phosphorylation. The elongation defect is explained by the loss of the elongation factors LEO1 and CDC73, part of PAF1 complex, and SPT6 from the newly-elongating pol II. Our results indicate that CDK12 is a general activator of pol II transcription elongation and indicate that it targets both Ser2 and Ser5 residues of the pol II CTD.
© The Author(s) 2020. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Zaborowska J., Egloff S., Murphy S.. The pol II CTD: new twists in the tail. Nat. Struct. Mol. Biol. 2016; 23:771–777. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
