

Repetitive Intermittent Hyperglycemia Drives the M1 Polarization and Inflammatory Responses in THP-1 Macrophages Through the Mechanism Involving the TLR4-IRF5 Pathway
- PMID: 32806763
- PMCID: PMC7463685
- DOI: 10.3390/cells9081892
Repetitive Intermittent Hyperglycemia Drives the M1 Polarization and Inflammatory Responses in THP-1 Macrophages Through the Mechanism Involving the TLR4-IRF5 Pathway
Abstract
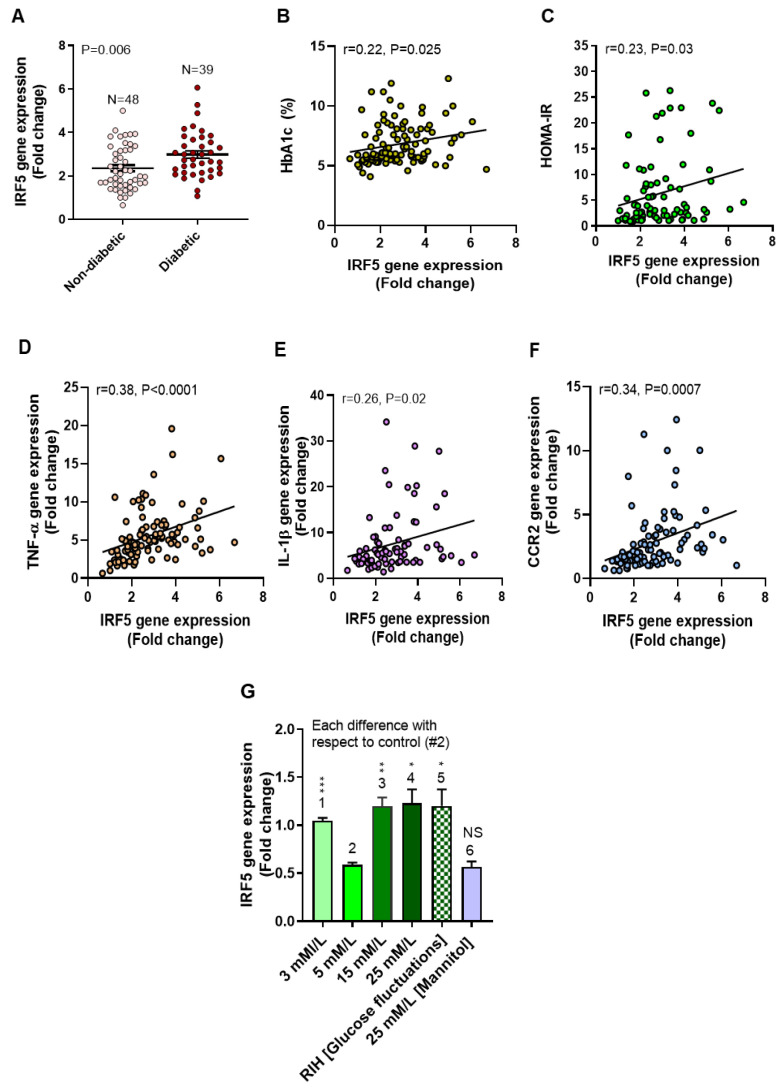
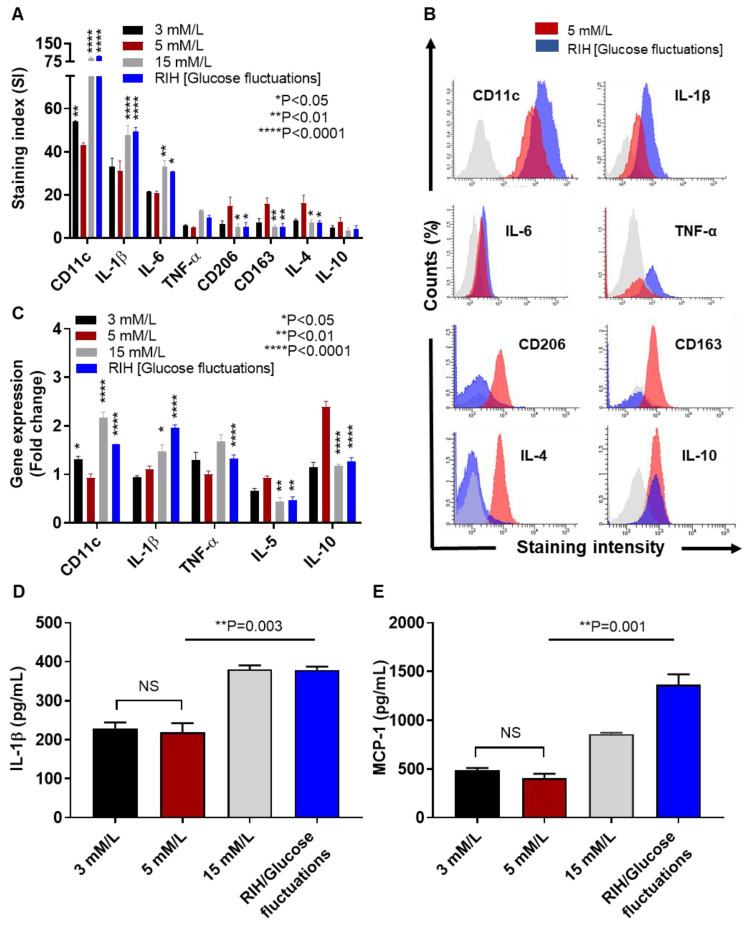
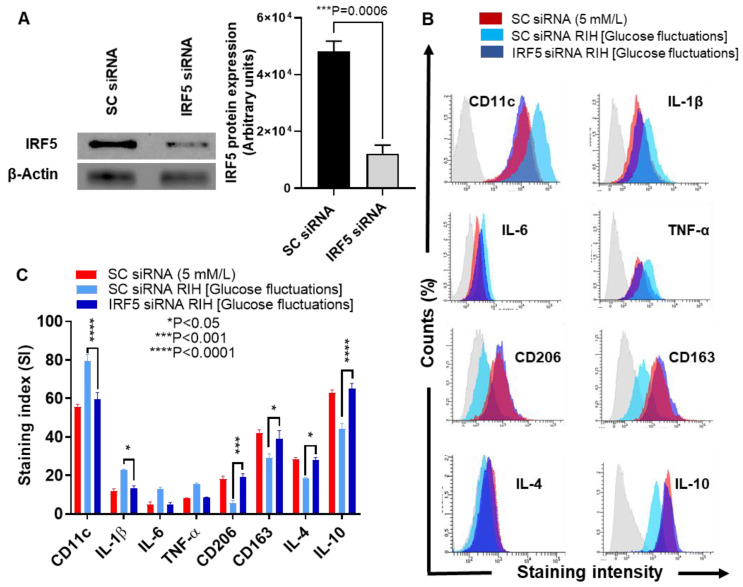
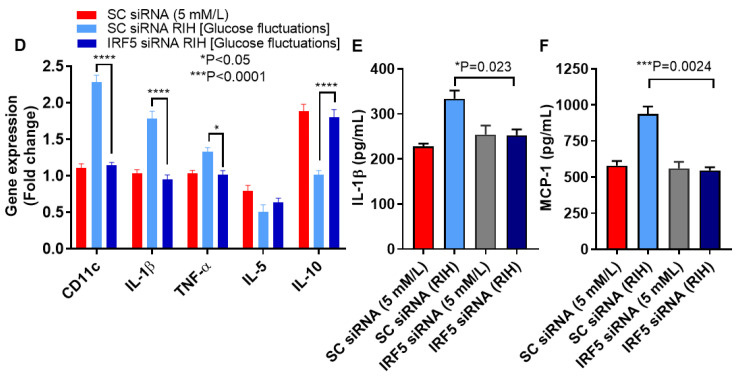
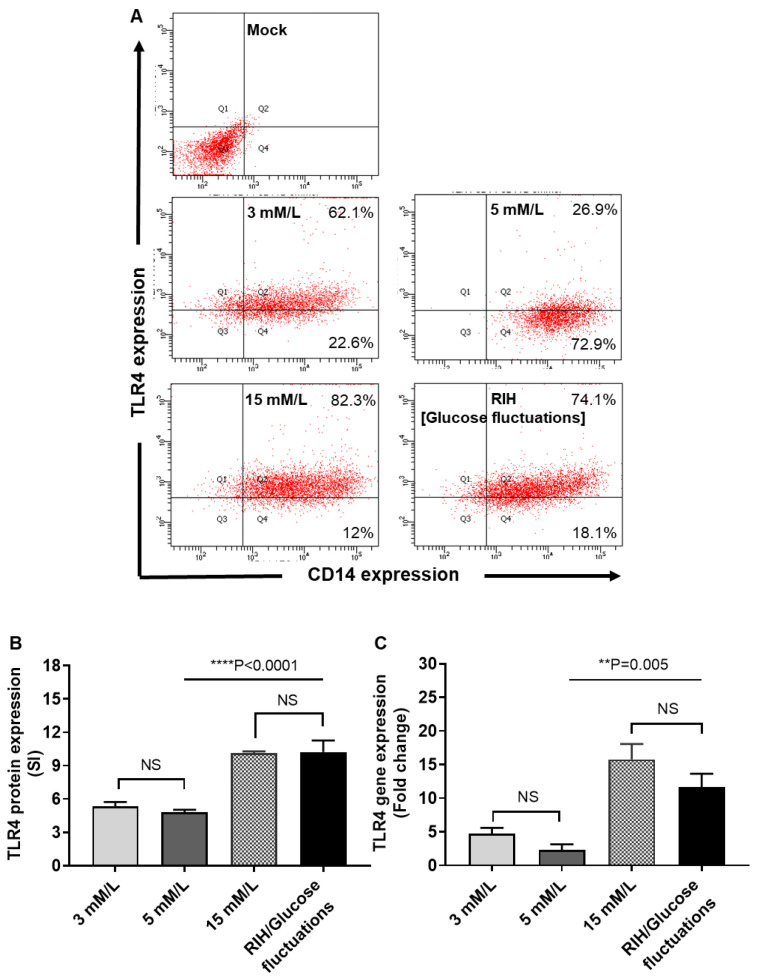
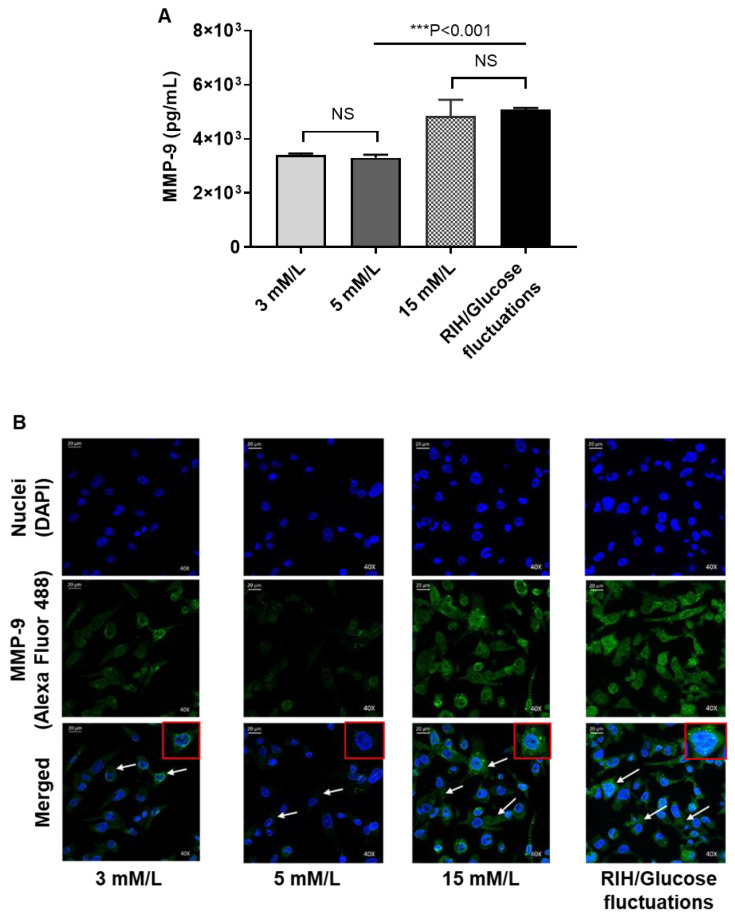
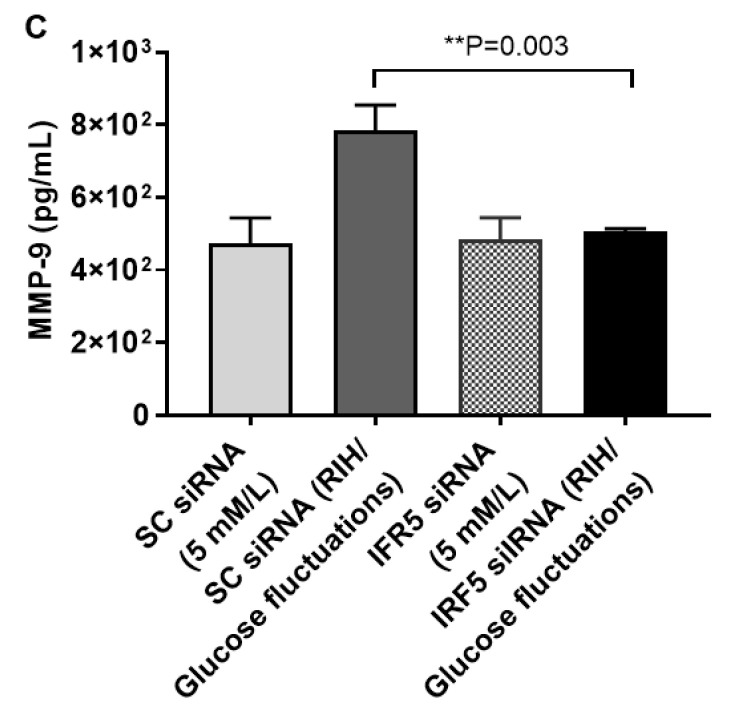
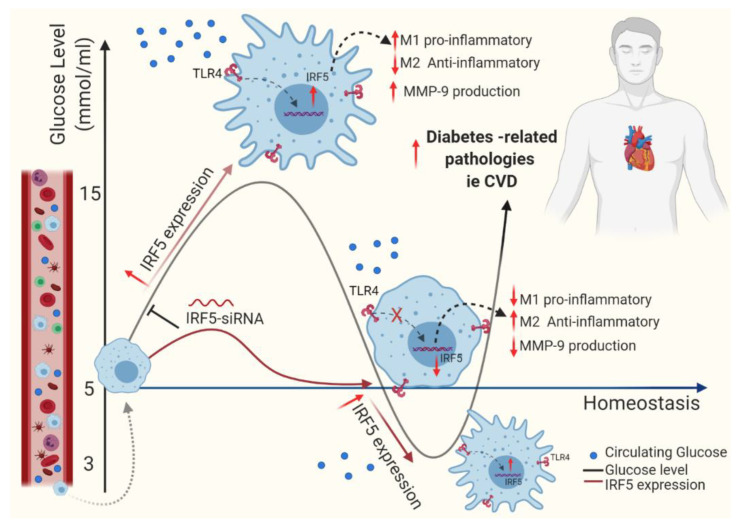
Repetitive intermittent hyperglycemia (RIH) is an independent risk factor for complications associated with type-2 diabetes (T2D). Glucose fluctuations commonly occur in T2D patients with poor glycemic control or following intensive therapy. Reducing blood glucose as well as glucose fluctuations is critical to the control of T2D and its macro-/microvascular complications. The interferon regulatory factor (IRF)-5 located downstream of the nutrient sensor toll-like receptor (TLR)-4, is emerging as a key metabolic regulator. It remains unclear how glucose fluctuations may alter the IRF5/TLR4 expression and inflammatory responses in monocytes/macrophages. To investigate this, first, we determined IRF5 gene expression by real-time qRT-PCR in the white adipose tissue samples from 39 T2D and 48 nondiabetic individuals. Next, we cultured THP-1 macrophages in hypo- and hyperglycemic conditions and compared, at the protein and transcription levels, the expressions of IRF5, TLR4, and M1/M2 polarization profile and inflammatory markers against control (normoglycemia). Protein expression was assessed using flow cytometry, ELISA, Western blotting, and/or confocal microscopy. IRF5 silencing was achieved by small interfering RNA (siRNA) transfection. The data show that adipose IRF5 gene expression was higher in T2D than nondiabetic counterparts (P = 0.006), which correlated with glycated hemoglobin (HbA1c) (r = 0.47/P < 0.001), homeostatic model assessment of insulin resistance (HOMA-IR) (r = 0.23/P = 0.03), tumor necrosis factor (TNF)-α (r = 0.56/P < 0.0001), interleukin (IL)-1β (r = 0.40/P = 0.0009), and C-C motif chemokine receptor (CCR)-2 (r = 0.49/P < 0.001) expression. IRF5 expression in macrophages was induced/upregulated (P < 0.05) by hypoglycemia (3 mM/L), persistent hyperglycemia (15 mM/L-25 mM/L), and RIH/glucose fluctuations (3-15 mM/L) as compared to normoglycemia (5 mM/L). RIH/glucose fluctuations also induced M1 polarization and an inflammatory profile (CD11c, IL-1β, TNF-α, IL-6, and monocyte chemoattractant protein (MCP)-1) in macrophages. RIH/glucose fluctuations also drove the expression of matrix metalloproteinase (MMP)-9 (P < 0.001), which is a known marker for cardiovascular complication in T2D patients. Notably, all these changes were counteracted by IRF5 silencing in macrophages. In conclusion, RIH/glucose fluctuations promote the M1 polarization and inflammatory responses in macrophages via the mechanism involving TLR4-IRF5 pathway, which may have significance for metabolic inflammation.
Keywords: IRF5; TLR4; glucose fluctuations; macrophages; metabolic inflammation; repetitive intermittent hyperglycemia; type-2 diabetes.
Conflict of interest statement
The authors declare that there are no conflicts of interest involved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
