

T-cells produce acidic niches in lymph nodes to suppress their own effector functions
- PMID: 32807791
- PMCID: PMC7431837
- DOI: 10.1038/s41467-020-17756-7
T-cells produce acidic niches in lymph nodes to suppress their own effector functions
Abstract
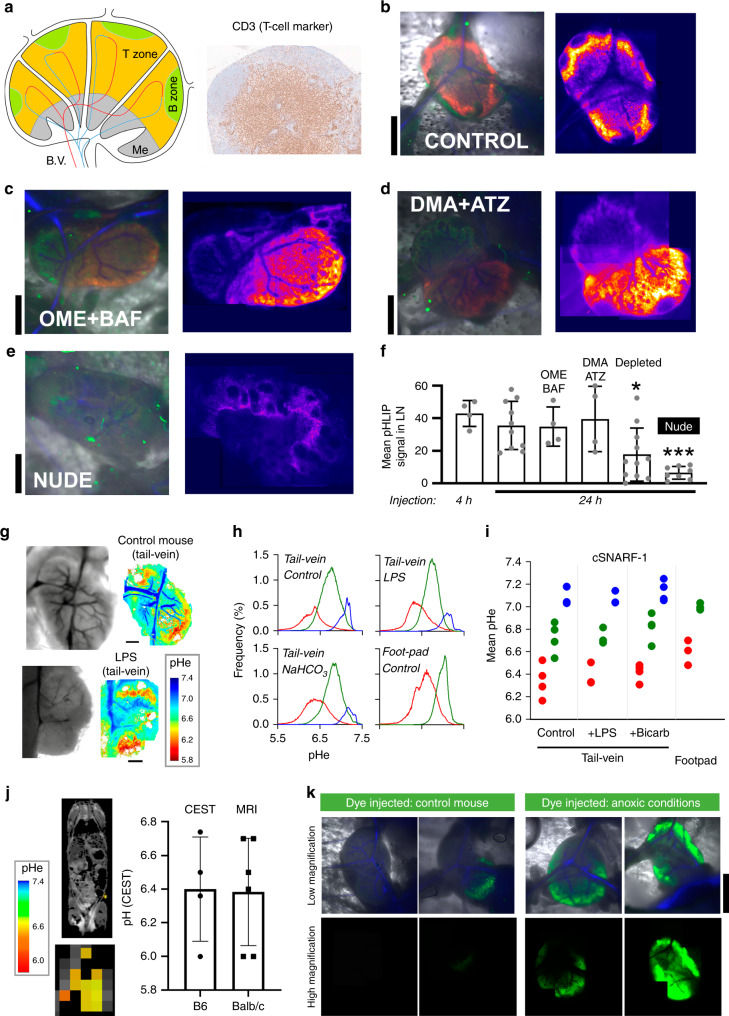
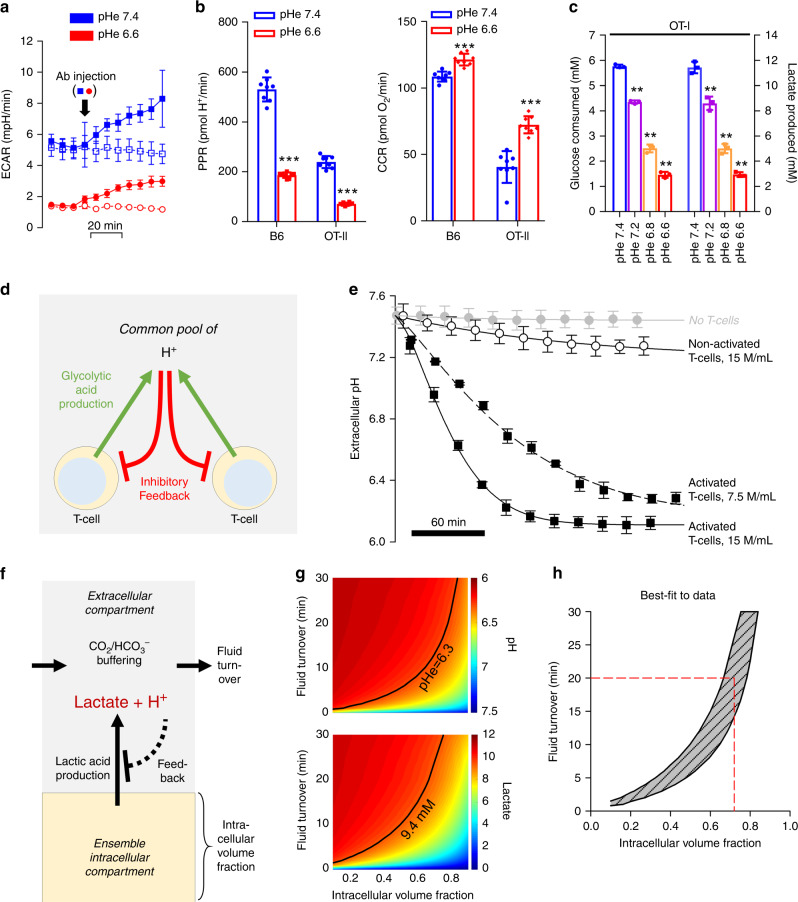
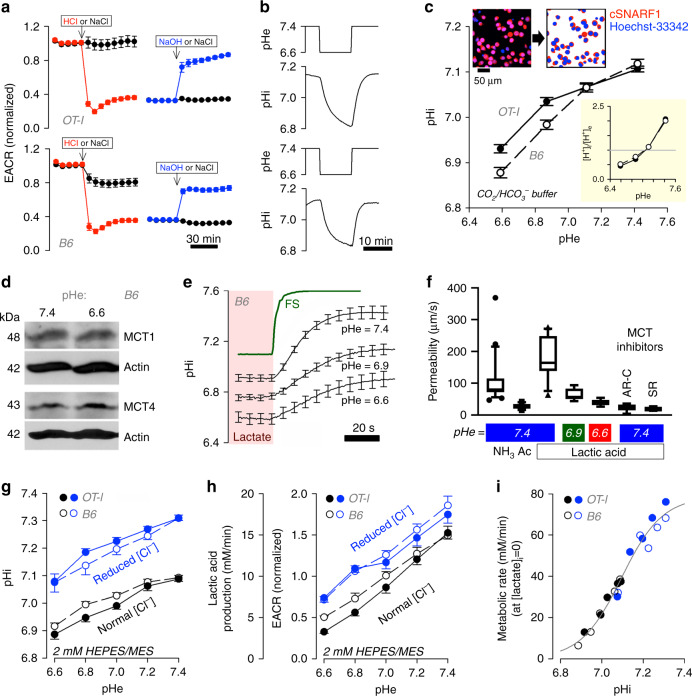
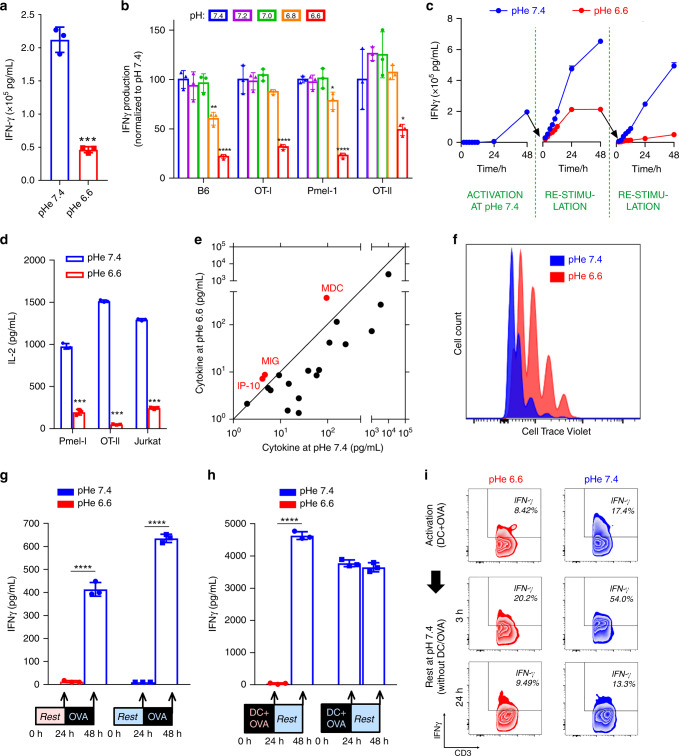
The acidic pH of tumors profoundly inhibits effector functions of activated CD8 + T-cells. We hypothesize that this is a physiological process in immune regulation, and that it occurs within lymph nodes (LNs), which are likely acidic because of low convective flow and high glucose metabolism. Here we show by in vivo fluorescence and MR imaging, that LN paracortical zones are profoundly acidic. These acidic niches are absent in athymic Nu/Nu and lymphodepleted mice, implicating T-cells in the acidifying process. T-cell glycolysis is inhibited at the low pH observed in LNs. We show that this is due to acid inhibition of monocarboxylate transporters (MCTs), resulting in a negative feedback on glycolytic rate. Importantly, we demonstrate that this acid pH does not hinder initial activation of naïve T-cells by dendritic cells. Thus, we describe an acidic niche within the immune system, and demonstrate its physiological role in regulating T-cell activation.
Conflict of interest statement
R.J.G. has research support form Helix Biopharma, who makes acid pH targeting agents. These agents were not used in the current work. O.A.A. and Y.K.R. are founders of pHLIP, Inc. They have shares in the company, but the company did not fund any part of the work reported in this paper, which was carried out in their academic laboratories. No other authors report competing interests.
Figures
Similar articles
-
Intratumoral interleukin-21 increases antitumor immunity, tumor-infiltrating CD8+ T-cell density and activity, and enlarges draining lymph nodes.J Immunother. 2010 Apr;33(3):236-49. doi: 10.1097/CJI.0b013e3181c0c1cb. J Immunother. 2010. PMID: 20445344
-
Acute and chronic B cell depletion disrupts CD4+ and CD8+ T cell homeostasis and expansion during acute viral infection in mice.J Immunol. 2014 Jul 15;193(2):746-56. doi: 10.4049/jimmunol.1302848. Epub 2014 Jun 13. J Immunol. 2014. PMID: 24928986 Free PMC article.
-
Organ-dependent in vivo priming of naive CD4+, but not CD8+, T cells by plasmacytoid dendritic cells.J Exp Med. 2007 Aug 6;204(8):1923-33. doi: 10.1084/jem.20062373. Epub 2007 Jul 23. J Exp Med. 2007. PMID: 17646404 Free PMC article.
-
The Chemoattractant Receptor Ebi2 Drives Intranodal Naive CD4+ T Cell Peripheralization to Promote Effective Adaptive Immunity.Immunity. 2019 May 21;50(5):1188-1201.e6. doi: 10.1016/j.immuni.2019.04.001. Epub 2019 Apr 30. Immunity. 2019. PMID: 31053504
-
CD103- and CD103+ bronchial lymph node dendritic cells are specialized in presenting and cross-presenting innocuous antigen to CD4+ and CD8+ T cells.J Immunol. 2007 Jun 1;178(11):6861-6. doi: 10.4049/jimmunol.178.11.6861. J Immunol. 2007. PMID: 17513734
Cited by
-
Overcoming current challenges to T-cell receptor therapy via metabolic targeting to increase antitumor efficacy, durability, and tolerability.Front Immunol. 2022 Nov 21;13:1056622. doi: 10.3389/fimmu.2022.1056622. eCollection 2022. Front Immunol. 2022. PMID: 36479131 Free PMC article. Review.
-
Multicolor Light-Induced Immune Activation via Polymer Photocaged Cytokines.Biomacromolecules. 2023 Mar 13;24(3):1164-1172. doi: 10.1021/acs.biomac.2c01207. Epub 2023 Feb 6. Biomacromolecules. 2023. PMID: 36745712 Free PMC article.
-
Tumor-draining lymph nodes: At the crossroads of metastasis and immunity.Sci Immunol. 2021 Sep 10;6(63):eabg3551. doi: 10.1126/sciimmunol.abg3551. Epub 2021 Sep 3. Sci Immunol. 2021. PMID: 34516744 Free PMC article. Review.
-
B Cells and IL-21-Producing Follicular Helper T Cells Cooperate to Determine the Dynamic Alterations of Premetastatic Tumor Draining Lymph Nodes of Breast Cancer.Research (Wash D C). 2024 Mar 29;7:0346. doi: 10.34133/research.0346. eCollection 2024. Research (Wash D C). 2024. PMID: 38559676 Free PMC article.
-
A review of lactate-lactylation in malignancy: its potential in immunotherapy.Front Immunol. 2024 May 8;15:1384948. doi: 10.3389/fimmu.2024.1384948. eCollection 2024. Front Immunol. 2024. PMID: 38779665 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
