

Methylglyoxal inhibits nuclear division through alterations in vacuolar morphology and accumulation of Atg18 on the vacuolar membrane in Saccharomyces cerevisiae
- PMID: 32807835
- PMCID: PMC7431575
- DOI: 10.1038/s41598-020-70802-8
Methylglyoxal inhibits nuclear division through alterations in vacuolar morphology and accumulation of Atg18 on the vacuolar membrane in Saccharomyces cerevisiae
Abstract
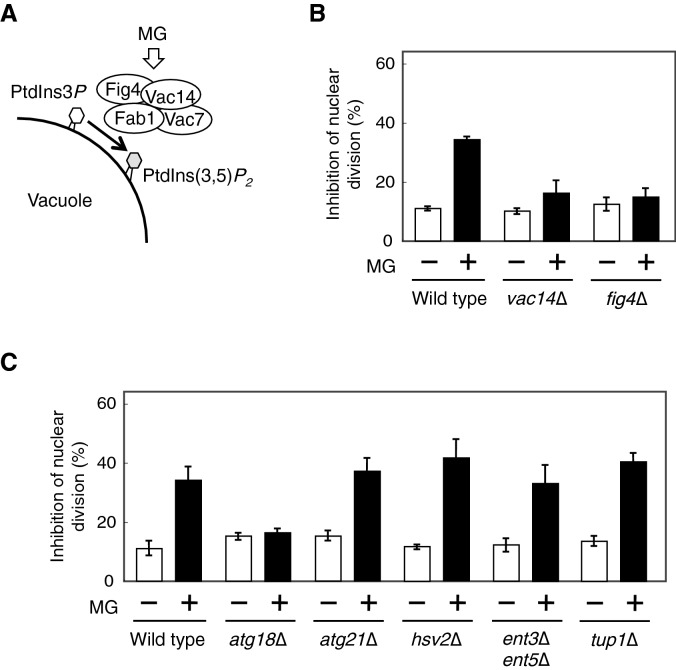
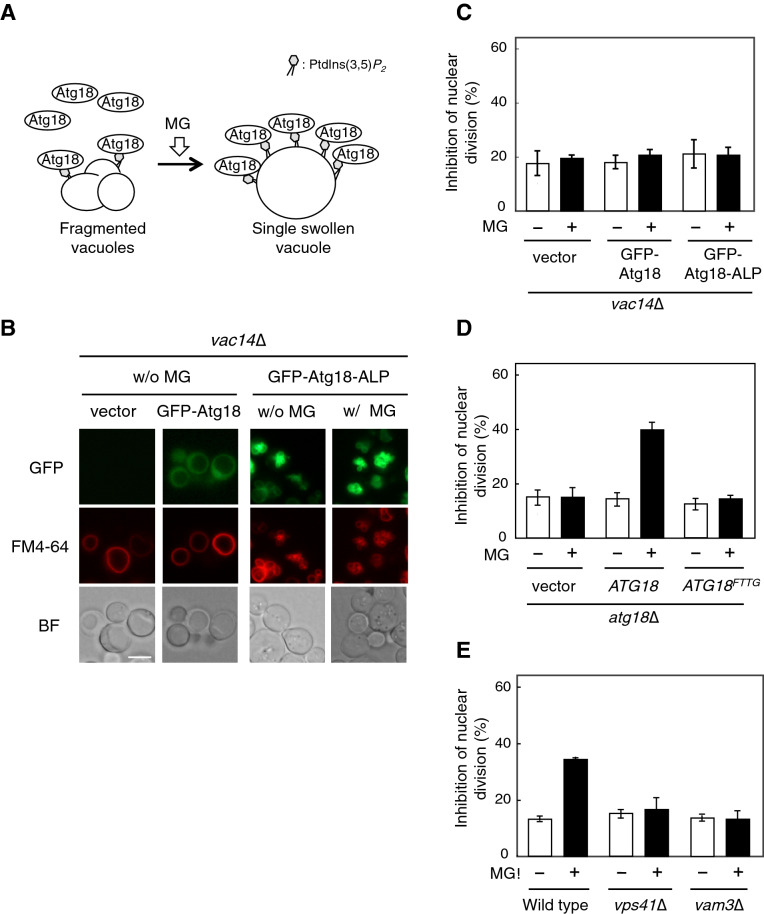
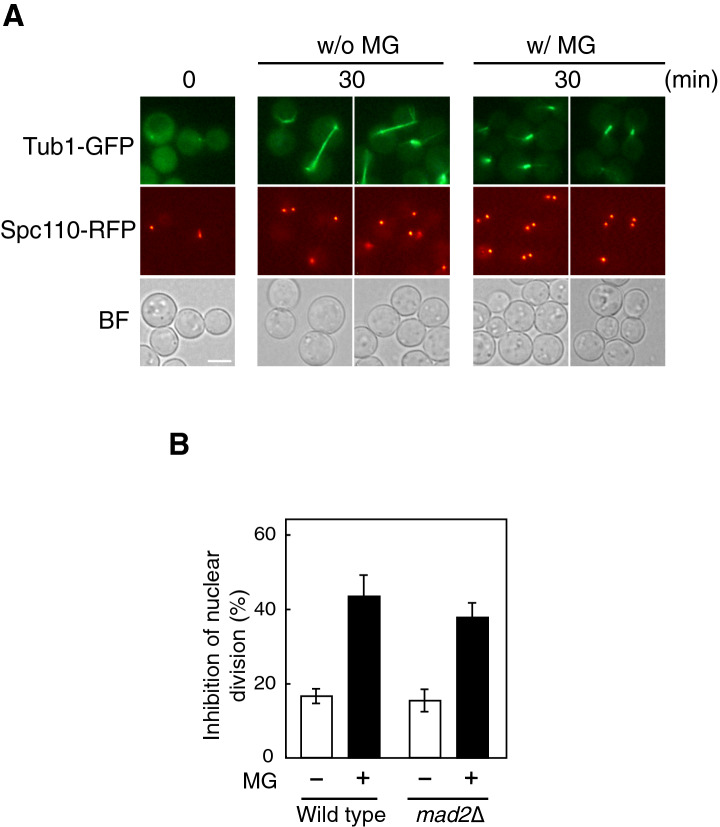
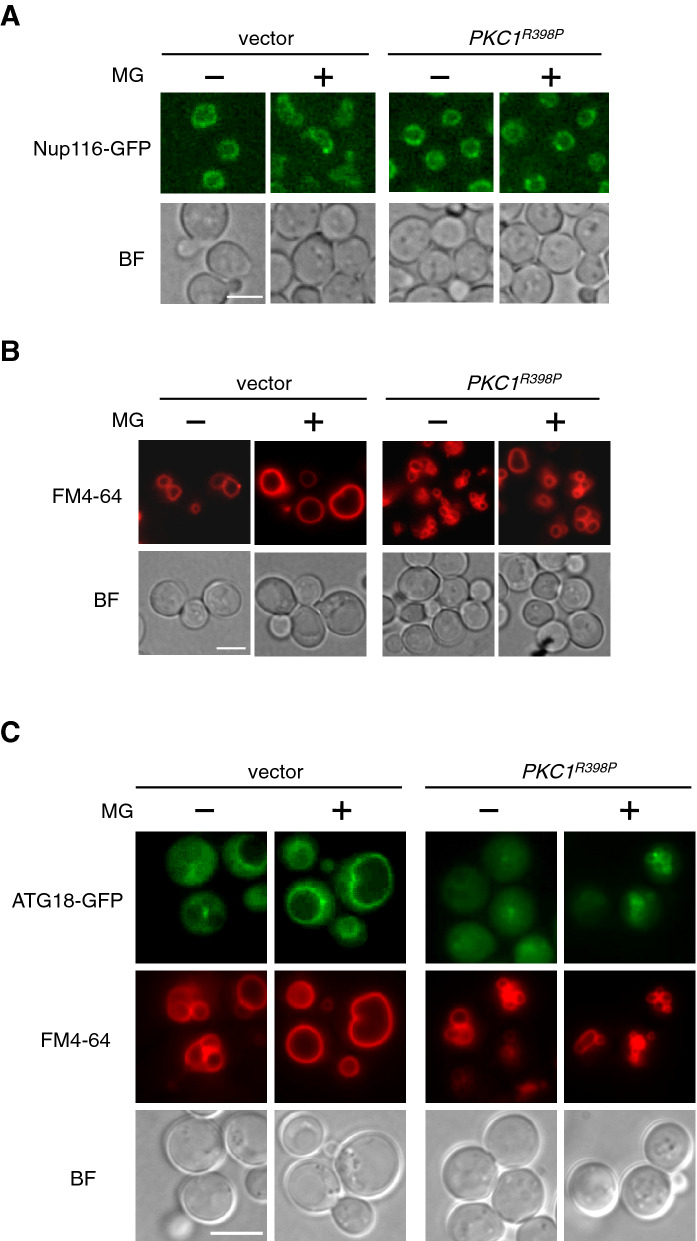
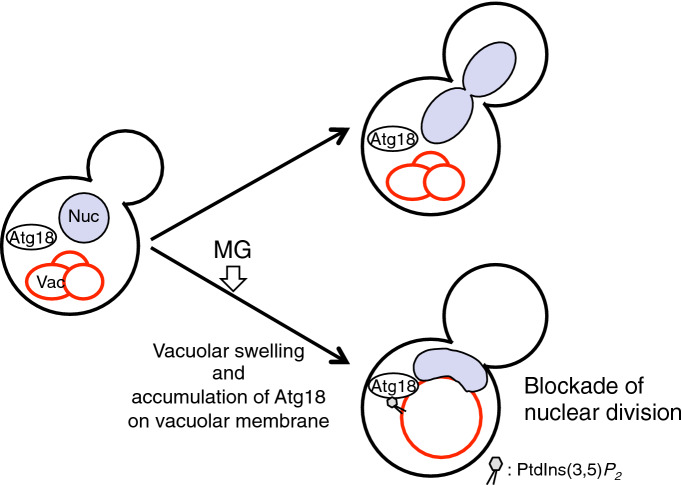
Methylglyoxal (MG) is a natural metabolite derived from glycolysis, and it inhibits the growth of cells in all kinds of organisms. We recently reported that MG inhibits nuclear division in Saccharomyces cerevisiae. However, the mechanism by which MG blocks nuclear division remains unclear. Here, we show that increase in the levels of phosphatidylinositol 3,5-bisphosphate (PtdIns(3,5)P2) is crucial for the inhibitory effects of MG on nuclear division, and the deletion of PtdIns(3,5)P2-effector Atg18 alleviated the MG-mediated inhibitory effects. Previously, we reported that MG altered morphology of the vacuole to a single swelling form, where PtdIns(3,5)P2 accumulates. The changes in the vacuolar morphology were also needed by MG to exert its inhibitory effects on nuclear division. The known checkpoint machinery, including the spindle assembly checkpoint and morphological checkpoint, are not involved in the blockade of nuclear division by MG. Our results suggest that both the accumulation of Atg18 on the vacuolar membrane and alterations in vacuolar morphology are necessary for the MG-induced inhibition of nuclear division.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Atg18 regulates organelle morphology and Fab1 kinase activity independent of its membrane recruitment by phosphatidylinositol 3,5-bisphosphate.Mol Biol Cell. 2007 Nov;18(11):4232-44. doi: 10.1091/mbc.e07-04-0301. Epub 2007 Aug 15. Mol Biol Cell. 2007. PMID: 17699591 Free PMC article.
-
The Atg18-Atg2 complex is recruited to autophagic membranes via phosphatidylinositol 3-phosphate and exerts an essential function.J Biol Chem. 2008 Aug 29;283(35):23972-80. doi: 10.1074/jbc.M803180200. Epub 2008 Jun 27. J Biol Chem. 2008. PMID: 18586673 Free PMC article.
-
Atg18 lifts up from and lands on the vacuolar membrane mediated by phosphorylation of its propellers.Autophagy. 2013 Dec;9(12):2161-2. doi: 10.4161/auto.26379. Epub 2013 Oct 22. Autophagy. 2013. PMID: 24145716
-
Our FABulous VACation: a decade of phosphatidylinositol 3,5-bisphosphate.Biochem Soc Symp. 2007;(74):129-39. doi: 10.1042/BSS0740129. Biochem Soc Symp. 2007. PMID: 17233586 Review.
-
Are vacuolar dynamics crucial factors for plant cell division and differentiation?Plant Sci. 2024 Jul;344:112090. doi: 10.1016/j.plantsci.2024.112090. Epub 2024 Apr 16. Plant Sci. 2024. PMID: 38636812 Review.
Cited by
-
Activation of the cell wall integrity pathway negatively regulates TORC2-Ypk1/2 signaling through blocking eisosome disassembly in Saccharomyces cerevisiae.Commun Biol. 2024 Jun 11;7(1):722. doi: 10.1038/s42003-024-06411-2. Commun Biol. 2024. PMID: 38862688 Free PMC article.
-
Nuclear ingression of cytoplasmic bodies accompanies a boost in autophagy.Life Sci Alliance. 2022 May 13;5(9):e202101160. doi: 10.26508/lsa.202101160. Print 2022 Sep. Life Sci Alliance. 2022. PMID: 35568434 Free PMC article.
-
The signalling lipid PI3,5P2 is essential for timely mitotic exit.Open Biol. 2023 Sep;13(9):230125. doi: 10.1098/rsob.230125. Epub 2023 Sep 27. Open Biol. 2023. PMID: 37751887 Free PMC article.
-
Methylglyoxal Alone or Combined with Light-Emitting Diodes/Complex Electromagnetic Fields Represent an Effective Response to Microbial Chronic Wound Infections.Antibiotics (Basel). 2025 Apr 10;14(4):396. doi: 10.3390/antibiotics14040396. Antibiotics (Basel). 2025. PMID: 40298537 Free PMC article.
References
-
- Cavanaugh AM, Jaspersen SL. Big lessons from little yeast: Budding and fission yeast centrosome structure, duplication, and function. Annu. Rev. Genet. 2017;51:361–383. - PubMed
-
- Cid VJ, et al. Orchestrating the cell cycle in yeast: sequential localization of key mitotic regulators at the spindle pole and the bud neck. Microbiology. 2002;148:2647–2659. - PubMed
-
- Inoue Y, Kimura A. Methylglyoxal and regulation of its metabolism in microorganisms. Adv. Microb. Physiol. 1995;37:177–227. - PubMed
-
- Inoue Y, Maeta K, Nomura W. Glyoxalase system in yeasts: structure, function, and physiology. Semin. Cell. Dev. Biol. 2011;22:278–284. - PubMed
-
- Zemva J, et al. Effects of the reactive metabolite methylglyoxal on cellular signalling, insulin action and metabolism—What we know in mammals and what we can learn from yeast. Exp. Clin. Endocrinol. Diabetes. 2019;127:203–214. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
