

Pheromone components affect motivation and induce persistent modulation of associative learning and memory in honey bees
- PMID: 32807870
- PMCID: PMC7431541
- DOI: 10.1038/s42003-020-01183-x
Pheromone components affect motivation and induce persistent modulation of associative learning and memory in honey bees
Abstract
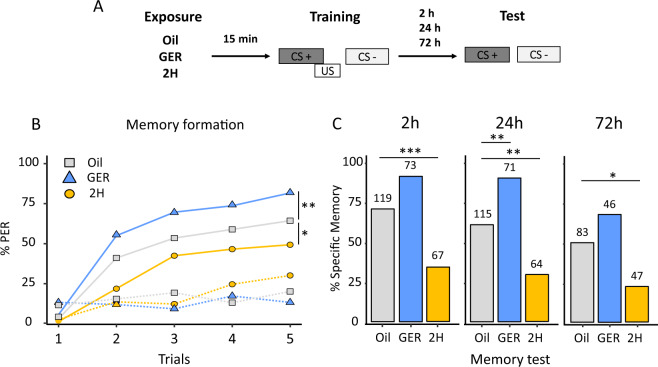
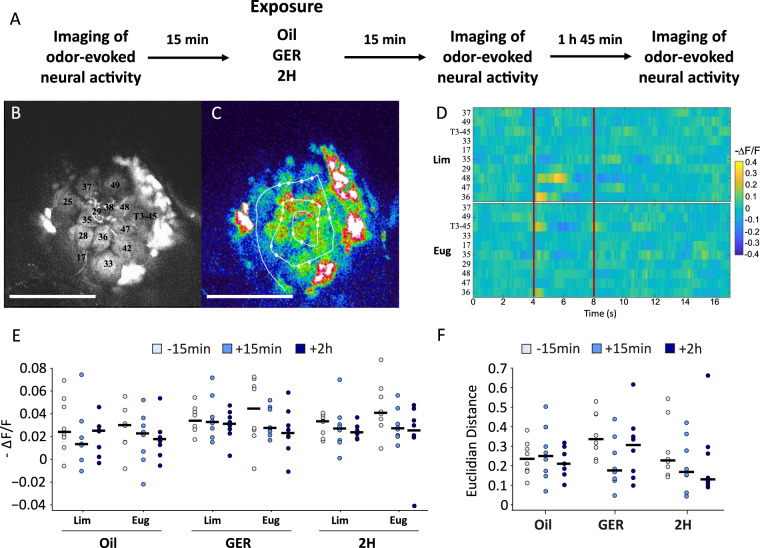
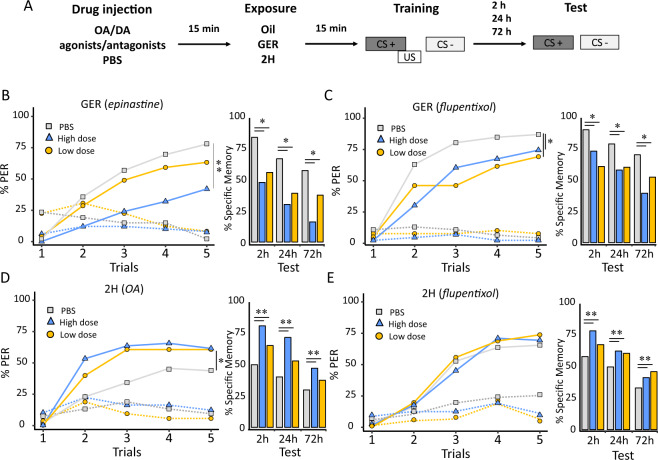
Since their discovery in insects, pheromones are considered as ubiquitous and stereotyped chemical messengers acting in intraspecific animal communication. Here we studied the effect of pheromones in a different context as we investigated their capacity to induce persistent modulations of associative learning and memory. We used honey bees, Apis mellifera, and combined olfactory conditioning and pheromone preexposure with disruption of neural activity and two-photon imaging of olfactory brain circuits, to characterize the effect of pheromones on olfactory learning and memory. Geraniol, an attractive pheromone component, and 2-heptanone, an aversive pheromone, improved and impaired, respectively, olfactory learning and memory via a durable modulation of appetitive motivation, which left odor processing unaffected. Consistently, interfering with aminergic circuits mediating appetitive motivation rescued or diminished the cognitive effects induced by pheromone components. We thus show that these chemical messengers act as important modulators of motivational processes and influence thereby animal cognition.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Karlson P, Luscher M. ‘Pheromones’: a new term for a class of biologically active substances. Nature. 1959;183:55–56. - PubMed
-
- Stowers L, Marton TF. What is a pheromone? Mammalian pheromones reconsidered. Neuron. 2005;46:699–702. - PubMed
-
- Coureaud G, et al. A pheromone that rapidly promotes learning in the newborn. Curr. Biol. 2006;16:1956–1961. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Medical
