

Polo kinase recruitment via the constitutive centromere-associated network at the kinetochore elevates centromeric RNA
- PMID: 32810142
- PMCID: PMC7455000
- DOI: 10.1371/journal.pgen.1008990
Polo kinase recruitment via the constitutive centromere-associated network at the kinetochore elevates centromeric RNA
Abstract
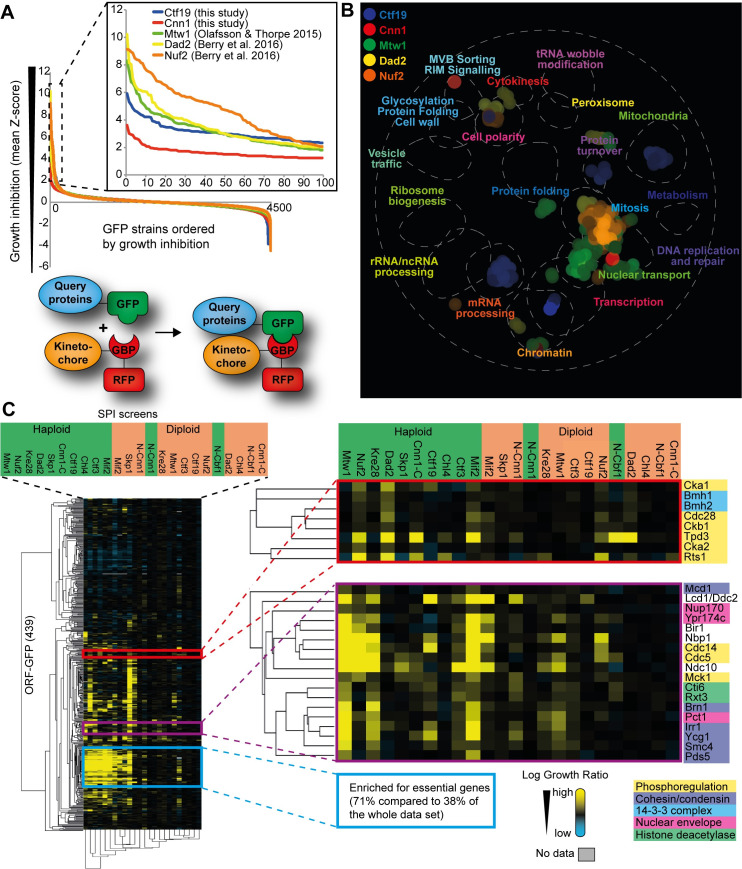
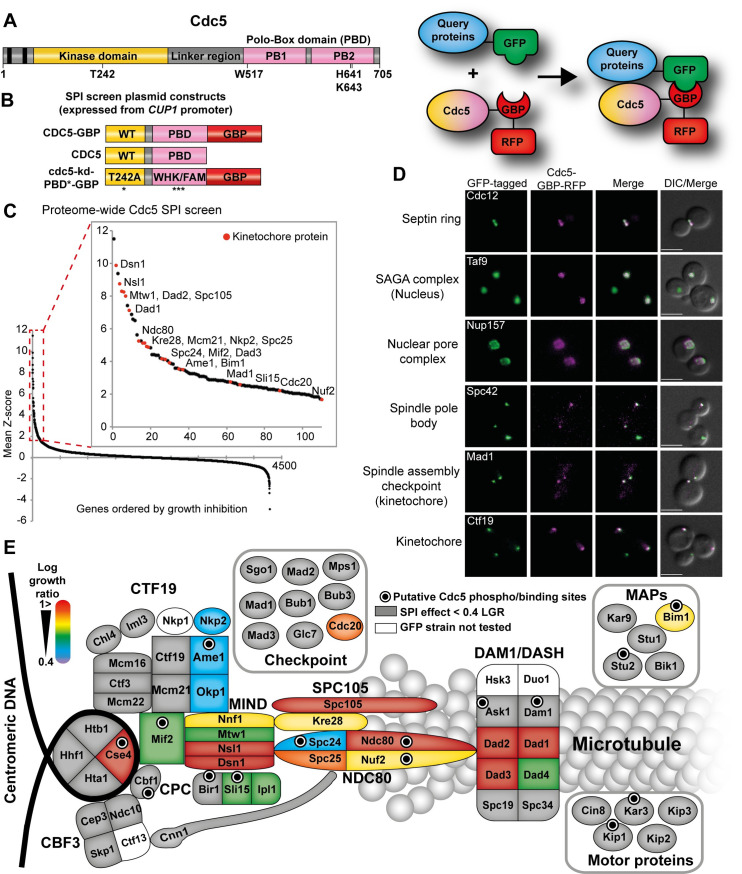
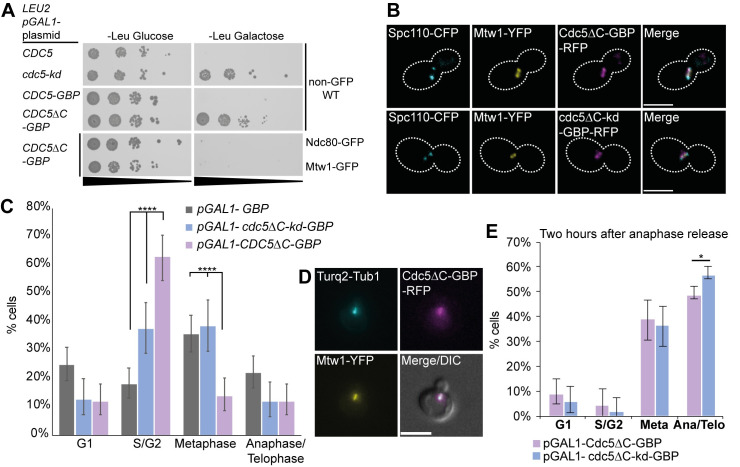
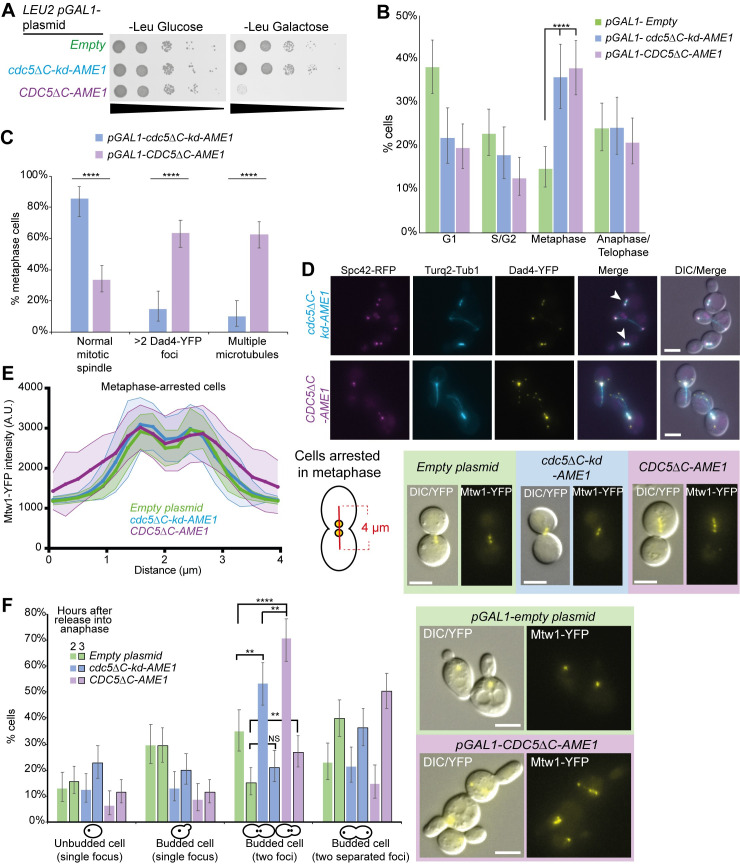
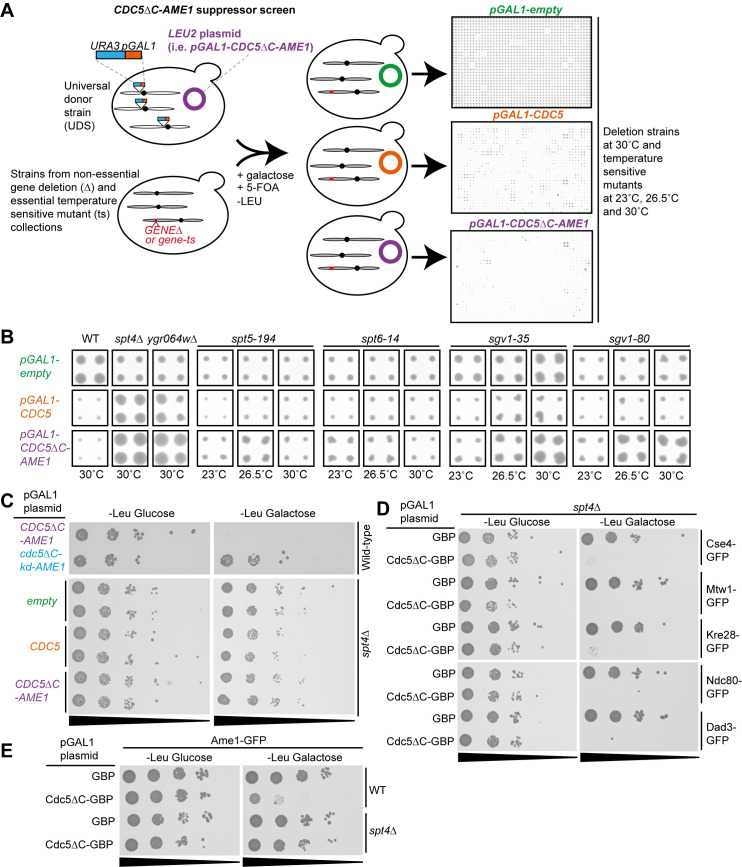
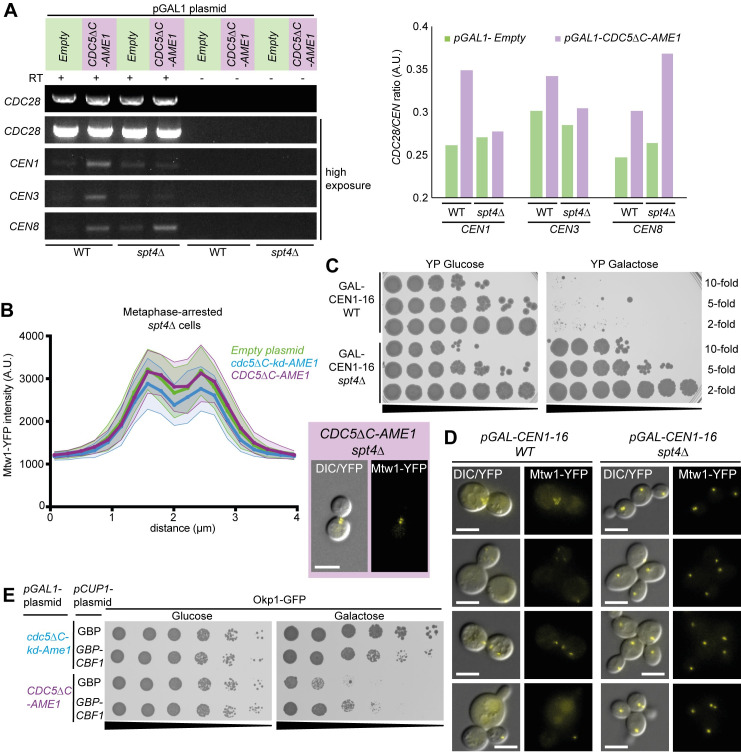
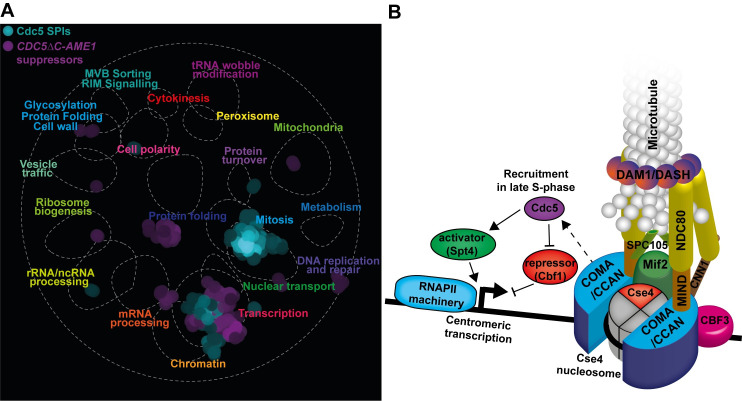
The kinetochore, a multi-protein complex assembled on centromeres, is essential to segregate chromosomes during cell division. Deficiencies in kinetochore function can lead to chromosomal instability and aneuploidy-a hallmark of cancer cells. Kinetochore function is controlled by recruitment of regulatory proteins, many of which have been documented, however their function often remains uncharacterized and many are yet to be identified. To identify candidates of kinetochore regulation we used a proteome-wide protein association strategy in budding yeast and detected many proteins that are involved in post-translational modifications such as kinases, phosphatases and histone modifiers. We focused on the Polo-like kinase, Cdc5, and interrogated which cellular components were sensitive to constitutive Cdc5 localization. The kinetochore is particularly sensitive to constitutive Cdc5 kinase activity. Targeting Cdc5 to different kinetochore subcomplexes produced diverse phenotypes, consistent with multiple distinct functions at the kinetochore. We show that targeting Cdc5 to the inner kinetochore, the constitutive centromere-associated network (CCAN), increases the levels of centromeric RNA via an SPT4 dependent mechanism.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
