

Stimulus-specific regulation of visual oddball differentiation in posterior parietal cortex
- PMID: 32811878
- PMCID: PMC7435179
- DOI: 10.1038/s41598-020-70448-6
Stimulus-specific regulation of visual oddball differentiation in posterior parietal cortex
Abstract
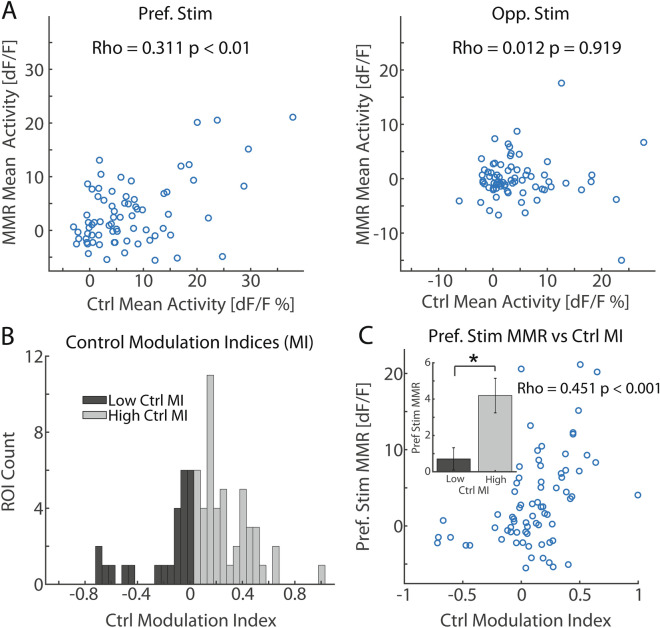
The frequency at which a stimulus is presented determines how it is interpreted. For example, a repeated image may be of less interest than an image that violates the prior sequence. This process involves integration of sensory information and internal representations of stimulus history, functions carried out in higher-order sensory areas such as the posterior parietal cortex (PPC). Thus far, there are few detailed reports investigating the single-neuron mechanisms for processing of stimulus presentation frequency in PPC. To address this gap in knowledge, we recorded PPC activity using 2-photon calcium imaging and electrophysiology during a visual oddball paradigm. Calcium imaging results reveal differentiation at the level of single neurons for frequent versus rare conditions which varied depending on whether the stimulus was preferred or non-preferred by the recorded neural population. Such differentiation of oddball conditions was mediated primarily by stimulus-independent adaptation in the frequent condition.
Conflict of interest statement
The authors declare no competing interests.
Figures
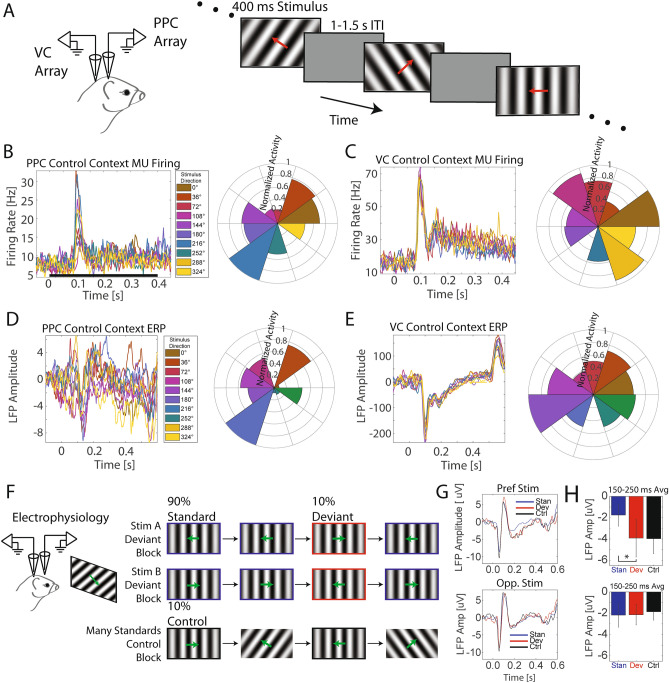
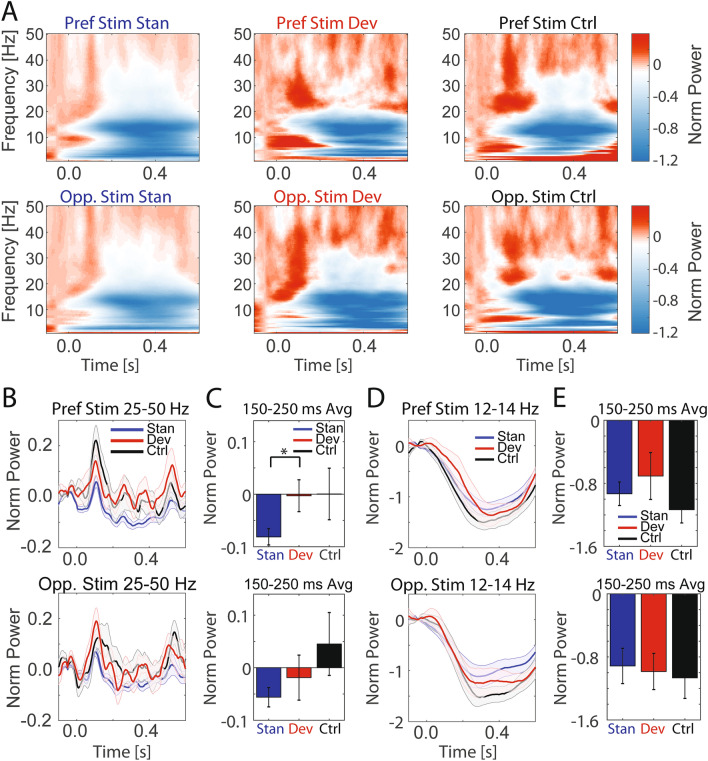
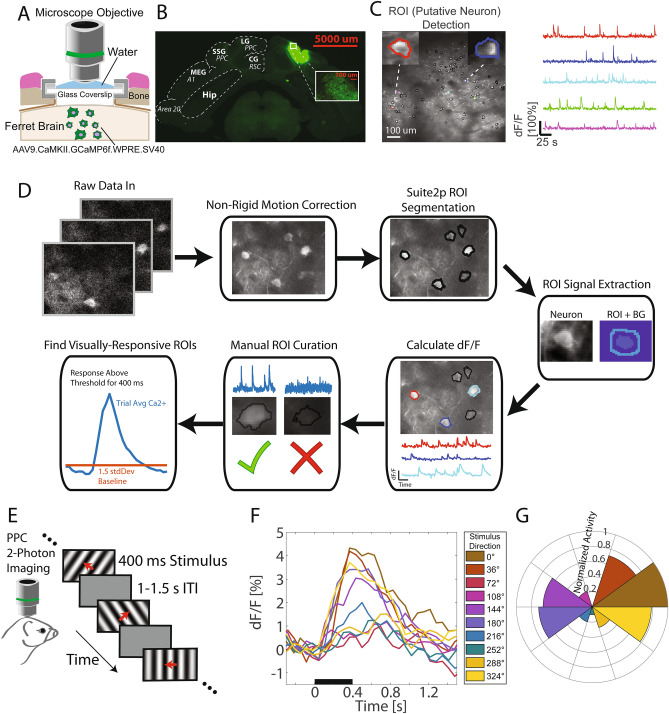
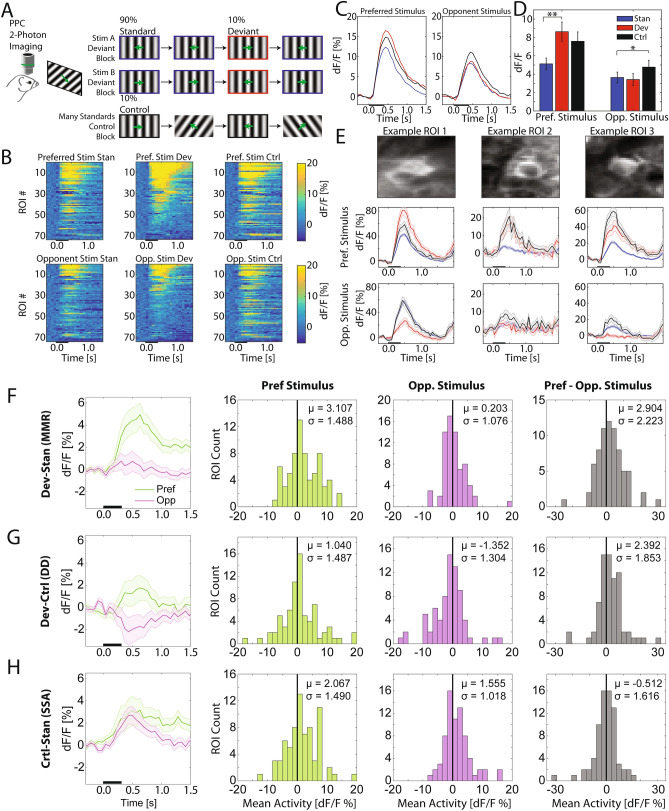
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
