

Interplay of Disorder and Sequence Specificity in the Formation of Stable Dynein-Dynactin Complexes
- PMID: 32814057
- PMCID: PMC7474177
- DOI: 10.1016/j.bpj.2020.07.023
Interplay of Disorder and Sequence Specificity in the Formation of Stable Dynein-Dynactin Complexes
Abstract
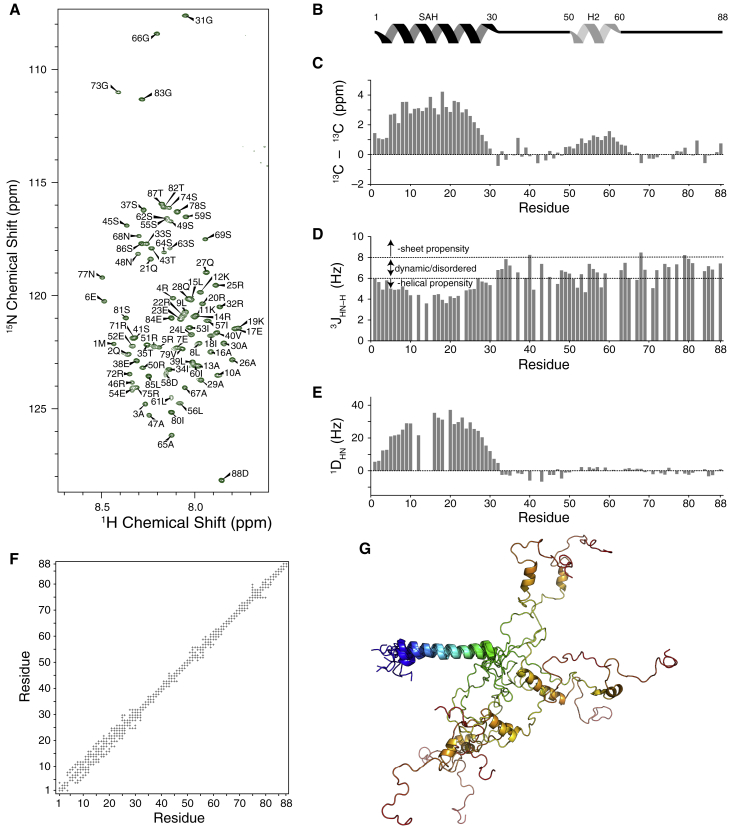
Cytoplasmic dynein is a eukaryotic motor protein complex that, along with its regulatory protein dynactin, is essential to the transport of organelles within cells. The interaction of dynein with dynactin is regulated by binding between the intermediate chain (IC) subunit of dynein and the p150Glued subunit of dynactin. Even though in the rat versions of these proteins this interaction primarily involves the single α-helix region at the N-terminus of the IC, in Drosophila and yeast ICs the removal of a nascent helix (H2) downstream of the single α-helix considerably diminishes IC-p150Glued complex stability. We find that for ICs from various species, there is a correlation between disorder in H2 and its contribution to binding affinity, and that sequence variations in H2 that do not change the level of disorder show similar binding behavior. Analysis of the structure and interactions of the IC from Chaetomium thermophilum demonstrates that the H2 region of C. thermophilum IC has a low helical propensity and establishes that H2 binds directly to the coiled-coil 1B (CC1B) domain of p150Glued, thus explaining why H2 is necessary for tight binding. Isothermal titration calorimetry, circular dichroism, and NMR studies of smaller CC1B constructs localize the region of CC1B most essential for a tight interaction with IC. These results suggest that it is the level of disorder in H2 of IC along with its charge, rather than sequence specificity, that underlie its importance in initiating tight IC-p150Glued complex formation. We speculate that the nascent H2 helix may provide conformational flexibility to initiate binding, whereas those species that have a fully folded H2 have co-opted an alternative mechanism for promoting p150Glued binding.
Copyright © 2020 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures








References
-
- Vallee R.B., McKenney R.J., Ori-McKenney K.M. Multiple modes of cytoplasmic dynein regulation. Nat. Cell Biol. 2012;14:224–230. - PubMed
-
- Kini A.R., Collins C.A. Modulation of cytoplasmic dynein ATPase activity by the accessory subunits. Cell Motil. Cytoskeleton. 2001;48:52–60. - PubMed
-
- Makokha M., Hare M., Barbar E. Interactions of cytoplasmic dynein light chains Tctex-1 and LC8 with the intermediate chain IC74. Biochemistry. 2002;41:4302–4311. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
