

A Plant Virus Ensures Viral Stability in the Hemolymph of Vector Insects through Suppressing Prophenoloxidase Activation
- PMID: 32817105
- PMCID: PMC7439478
- DOI: 10.1128/mBio.01453-20
A Plant Virus Ensures Viral Stability in the Hemolymph of Vector Insects through Suppressing Prophenoloxidase Activation
Abstract
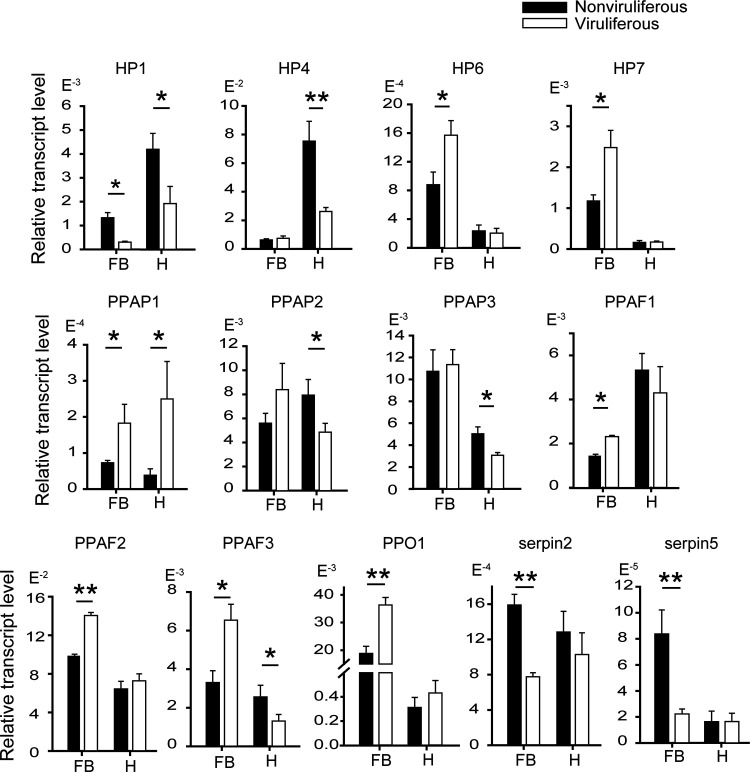
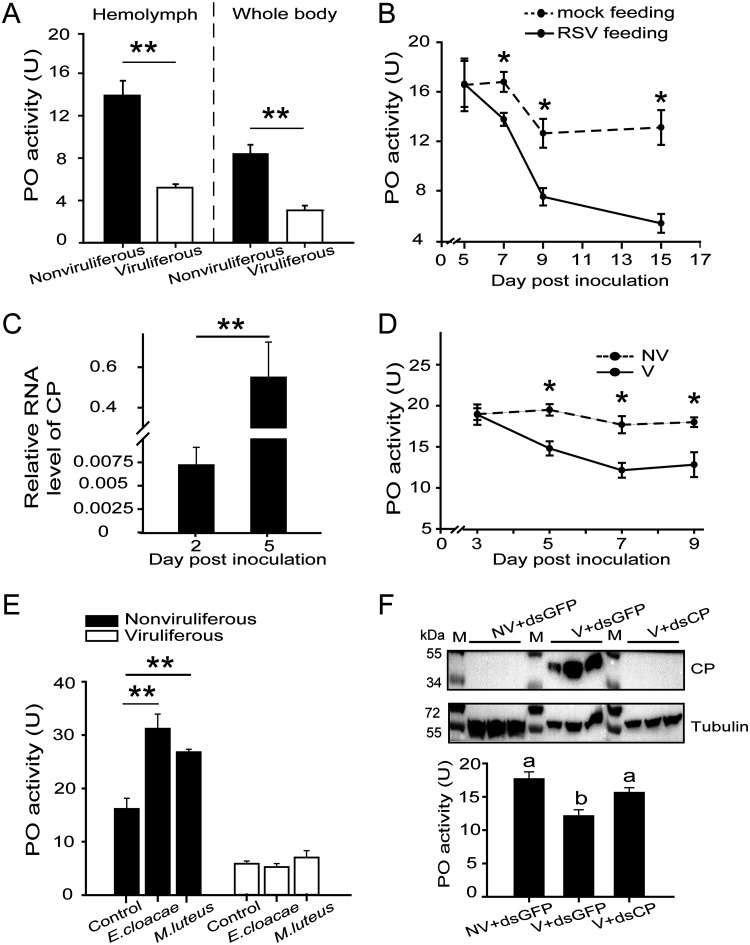
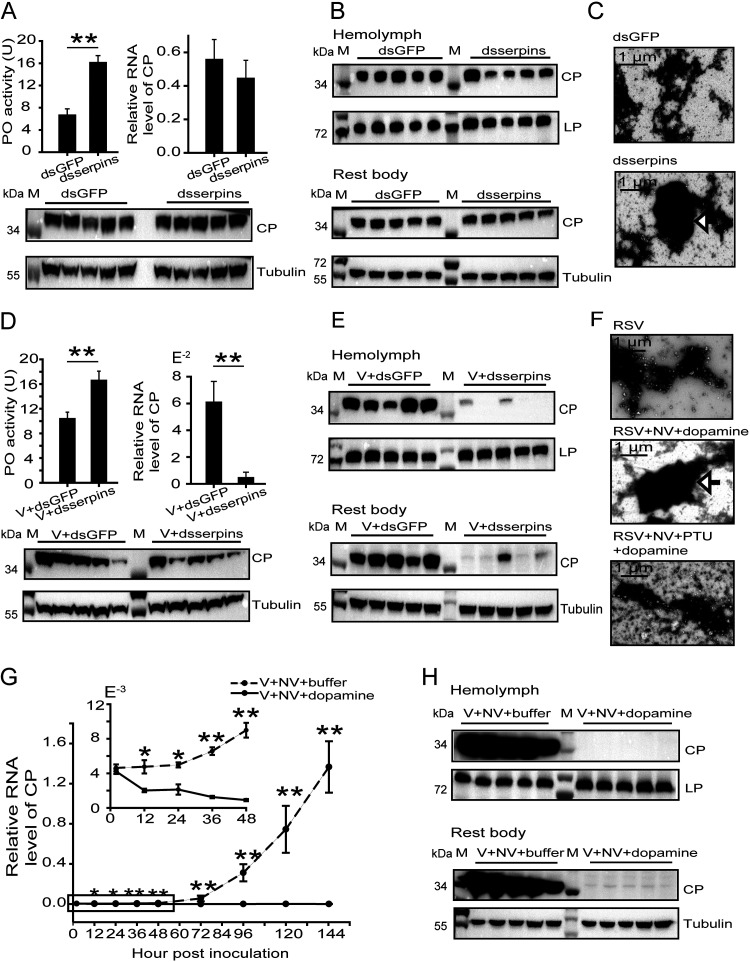
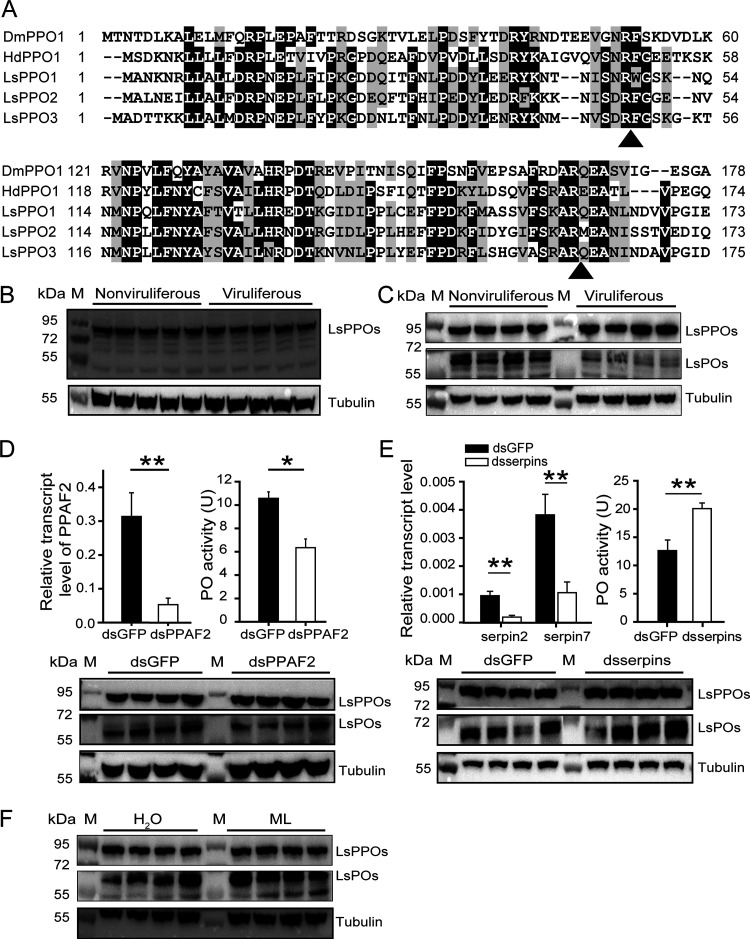
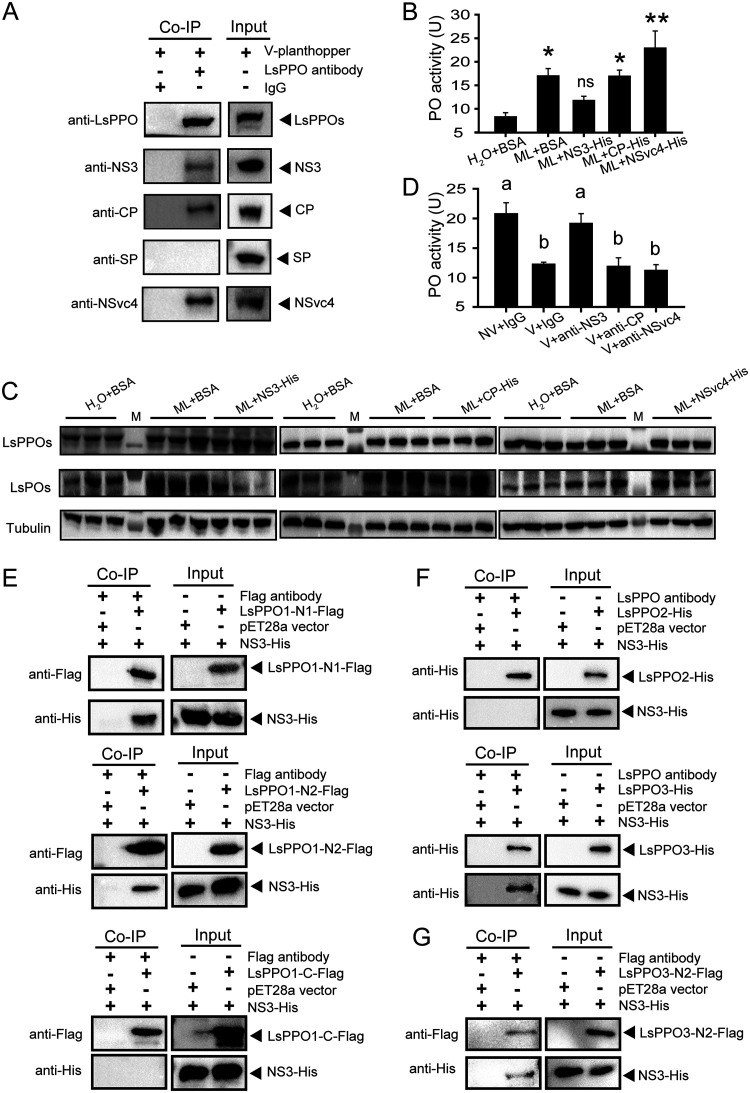
Most plant viruses require vector insects for transmission. Viral stability in the hemolymph of vector insects is a prerequisite for successful transmission of persistent plant viruses. However, knowledge of whether the proteolytic activation of prophenoloxidase (PPO) affects the stability of persistent plant viruses remains elusive. Here, we explored the interplay between rice stripe virus (RSV) and the PPO cascade of the vector small brown planthopper. Phenoloxidase (PO) activity was suppressed by RSV by approximately 60%. When the PPO cascade was activated, we found distinct melanization around RSV particles and serious damage to viral stability in the hemolymph. Viral suppression of PO activity was derived from obstruction of proteolytic cleavage of PPOs by binding of the viral nonstructural protein NS3. These results indicate that RSV attenuates the PPO response to ensure viral stability in the hemolymph of vector insects. Our research provides enlightening cues for controlling the transmission of vector-borne viruses.IMPORTANCE Large ratios of vector-borne plant viruses circulate in the hemolymph of their vector insects before entering the salivary glands to be transmitted to plants. The stability of virions in the hemolymph is vital in this process. Activation of the proteolytic prophenoloxidase (PPO) to produce active phenoloxidase (PO) is one of the major innate immune pathways in insect hemolymph. How a plant virus copes with the PPO immune reaction in its vector insect remains unclear. Here, we report that the PPO affects the stability of rice stripe virus (RSV), a notorious rice virus, in the hemolymph of a vector insect, the small brown planthopper. RSV suppresses PPO activation using viral nonstructural protein. Once the level of PO activity is elevated, RSV is melanized and eliminated from the hemolymph. Our work gives valuable clues for developing novel strategies for controlling the transmission of vector-borne plant viruses.
Keywords: NS3; hemolymph; plant virus; prophenoloxidase; rice stripe virus; small brown planthopper; vector insect.
Copyright © 2020 Chen et al.
Figures
Similar articles
-
A plant virus attenuates the Toll immune pathway by degradation of Pellino to facilitate viral infection in insect vectors.J Virol. 2025 May 20;99(5):e0002125. doi: 10.1128/jvi.00021-25. Epub 2025 Mar 31. J Virol. 2025. PMID: 40162791 Free PMC article.
-
Flotillin 2 Facilitates the Infection of a Plant Virus in the Gut of Insect Vector.J Virol. 2022 Apr 13;96(7):e0214021. doi: 10.1128/jvi.02140-21. Epub 2022 Mar 7. J Virol. 2022. PMID: 35254088 Free PMC article.
-
An insect cell line derived from the small brown planthopper supports replication of rice stripe virus, a tenuivirus.J Gen Virol. 2013 Jun;94(Pt 6):1421-1425. doi: 10.1099/vir.0.050104-0. Epub 2013 Mar 6. J Gen Virol. 2013. PMID: 23468422
-
Vector mediated transmission of persistently transmitted plant viruses.Curr Opin Virol. 2018 Feb;28:127-132. doi: 10.1016/j.coviro.2017.12.004. Epub 2018 Jan 4. Curr Opin Virol. 2018. PMID: 29306179 Review.
-
The small brown planthopper (Laodelphax striatellus) as a vector of the rice stripe virus.Arch Insect Biochem Physiol. 2023 Feb;112(2):e21992. doi: 10.1002/arch.21992. Epub 2022 Dec 27. Arch Insect Biochem Physiol. 2023. PMID: 36575628 Review.
Cited by
-
Regulation of RNA Interference Pathways in the Insect Vector Laodelphax striatellus by Viral Proteins of Rice Stripe Virus.Viruses. 2021 Aug 11;13(8):1591. doi: 10.3390/v13081591. Viruses. 2021. PMID: 34452456 Free PMC article.
-
A phytoplasma effector suppresses insect melanization immune response to promote pathogen persistent transmission.Sci Adv. 2025 Jan 31;11(5):eads9781. doi: 10.1126/sciadv.ads9781. Epub 2025 Jan 29. Sci Adv. 2025. PMID: 39879313 Free PMC article.
-
A plant virus attenuates the Toll immune pathway by degradation of Pellino to facilitate viral infection in insect vectors.J Virol. 2025 May 20;99(5):e0002125. doi: 10.1128/jvi.00021-25. Epub 2025 Mar 31. J Virol. 2025. PMID: 40162791 Free PMC article.
-
The nucleocapsid protein of rice stripe virus in cell nuclei of vector insect regulates viral replication.Protein Cell. 2022 May;13(5):360-378. doi: 10.1007/s13238-021-00822-1. Epub 2021 Mar 6. Protein Cell. 2022. PMID: 33675514 Free PMC article.
-
The Bunyavirales: The Plant-Infecting Counterparts.Viruses. 2021 May 6;13(5):842. doi: 10.3390/v13050842. Viruses. 2021. PMID: 34066457 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
