Estimating the genome-wide contribution of selection to temporal allele frequency change
- PMID: 32817464
- PMCID: PMC7456072
- DOI: 10.1073/pnas.1919039117
Estimating the genome-wide contribution of selection to temporal allele frequency change
Abstract
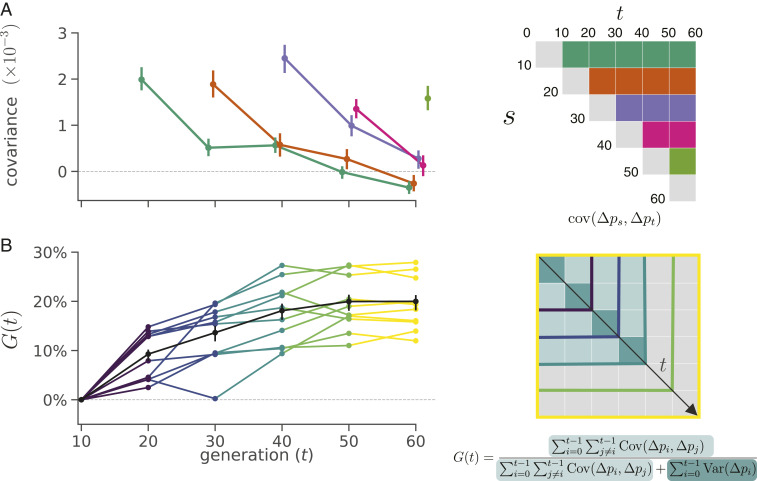
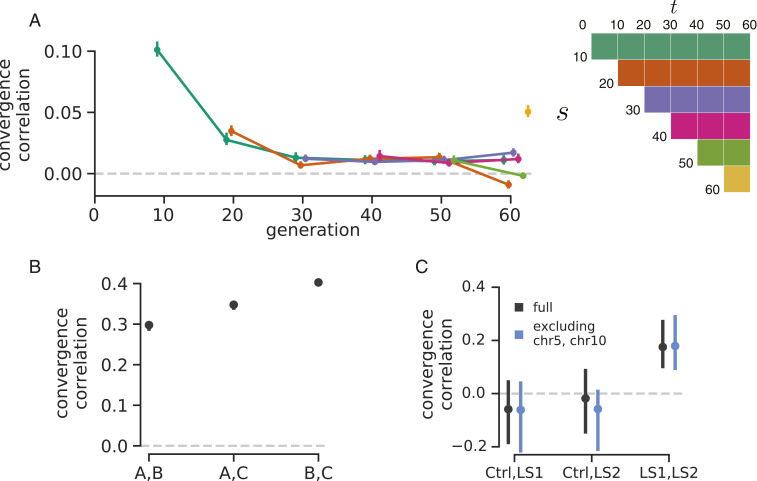
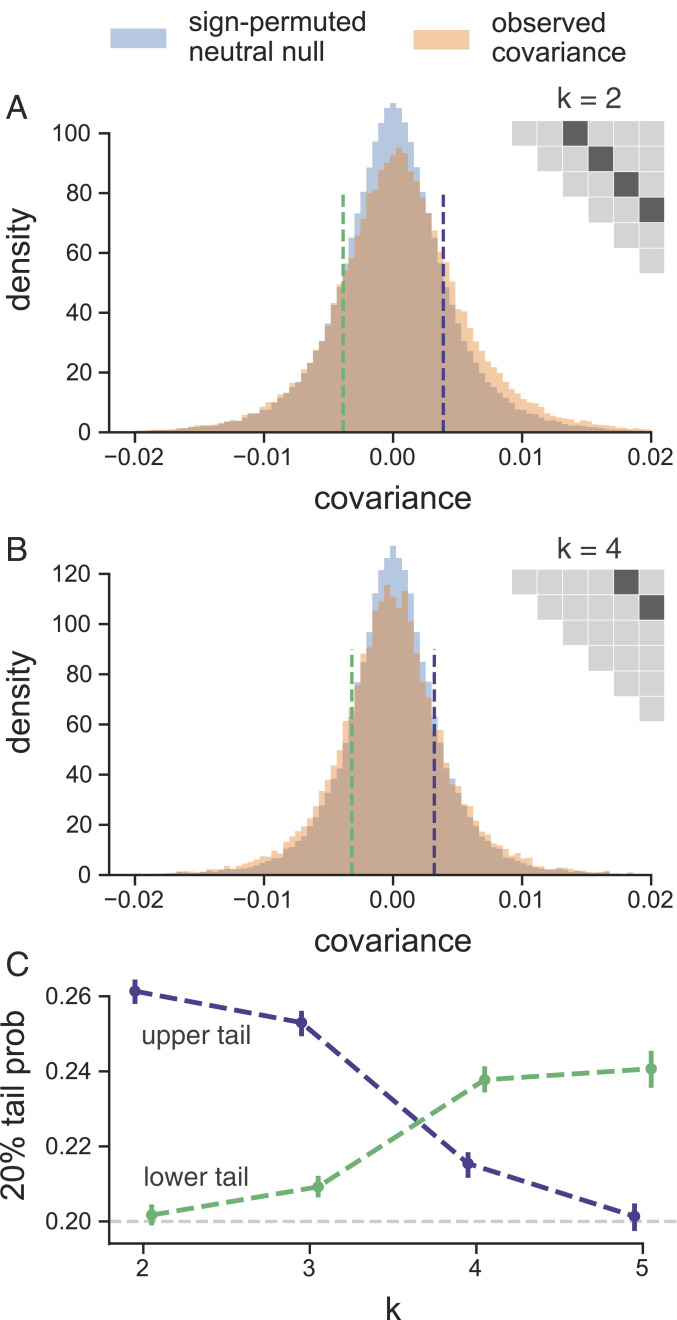
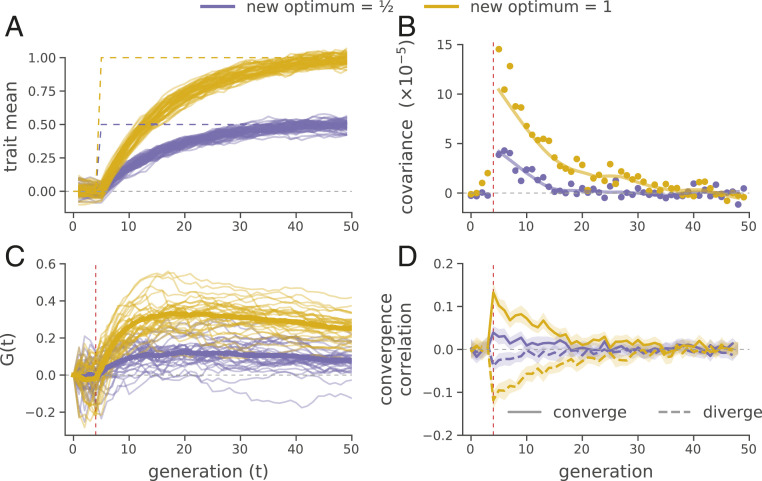
Rapid phenotypic adaptation is often observed in natural populations and selection experiments. However, detecting the genome-wide impact of this selection is difficult since adaptation often proceeds from standing variation and selection on polygenic traits, both of which may leave faint genomic signals indistinguishable from a noisy background of genetic drift. One promising signal comes from the genome-wide covariance between allele frequency changes observable from temporal genomic data (e.g., evolve-and-resequence studies). These temporal covariances reflect how heritable fitness variation in the population leads changes in allele frequencies at one time point to be predictive of the changes at later time points, as alleles are indirectly selected due to remaining associations with selected alleles. Since genetic drift does not lead to temporal covariance, we can use these covariances to estimate what fraction of the variation in allele frequency change through time is driven by linked selection. Here, we reanalyze three selection experiments to quantify the effects of linked selection over short timescales using covariance among time points and across replicates. We estimate that at least 17 to 37% of allele frequency change is driven by selection in these experiments. Against this background of positive genome-wide temporal covariances, we also identify signals of negative temporal covariance corresponding to reversals in the direction of selection for a reasonable proportion of loci over the time course of a selection experiment. Overall, we find that in the three studies we analyzed, linked selection has a large impact on short-term allele frequency dynamics that is readily distinguishable from genetic drift.
Keywords: adaptation; experimental evolution; linked selection.
Conflict of interest statement
The authors declare no competing interest.
Figures