

PRDM8 reveals aberrant DNA methylation in aging syndromes and is relevant for hematopoietic and neuronal differentiation
- PMID: 32819411
- PMCID: PMC7439574
- DOI: 10.1186/s13148-020-00914-5
PRDM8 reveals aberrant DNA methylation in aging syndromes and is relevant for hematopoietic and neuronal differentiation
Abstract
Background: Dyskeratosis congenita (DKC) and idiopathic aplastic anemia (AA) are bone marrow failure syndromes that share characteristics of premature aging with severe telomere attrition. Aging is also reflected by DNA methylation changes, which can be utilized to predict donor age. There is evidence that such epigenetic age predictions are accelerated in premature aging syndromes, but it is yet unclear how this is related to telomere length. DNA methylation analysis may support diagnosis of DKC and AA, which still remains a challenge for these rare diseases.
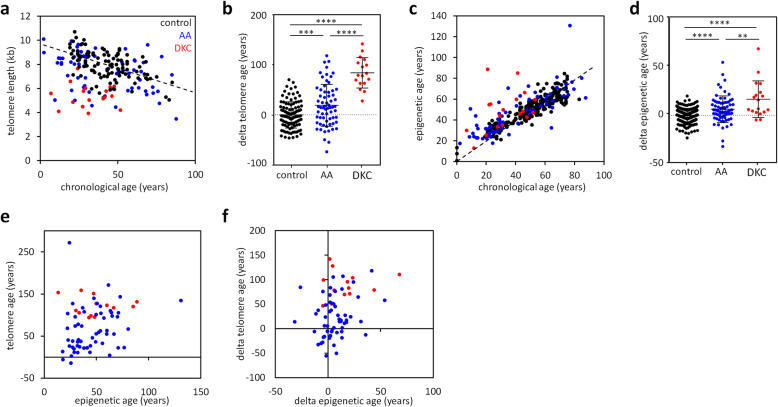
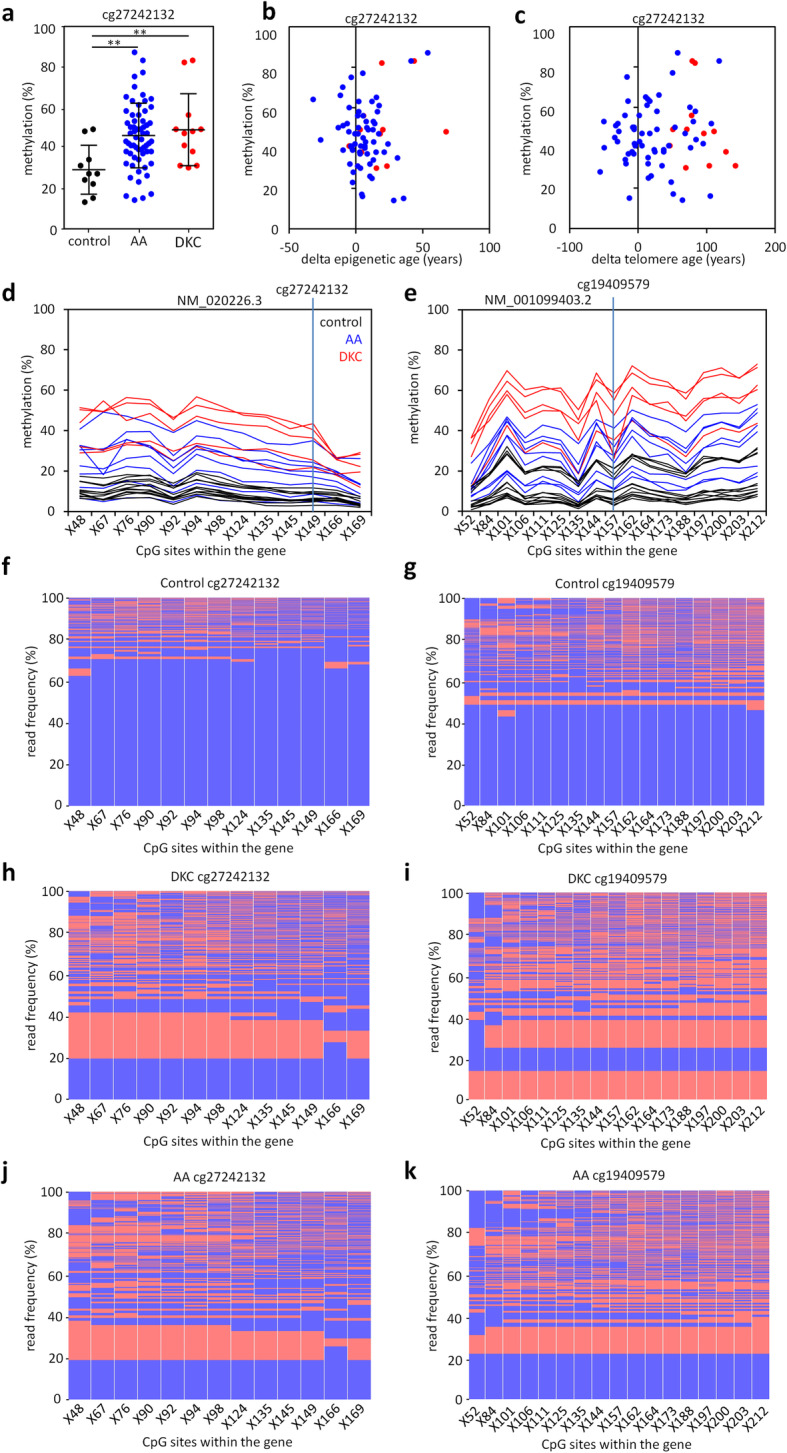
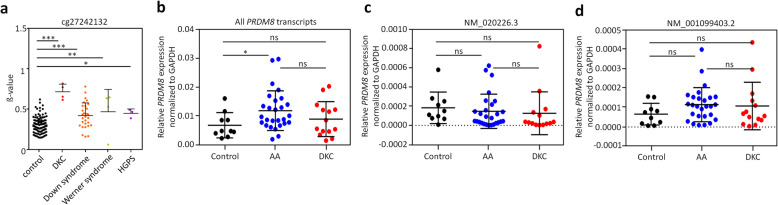
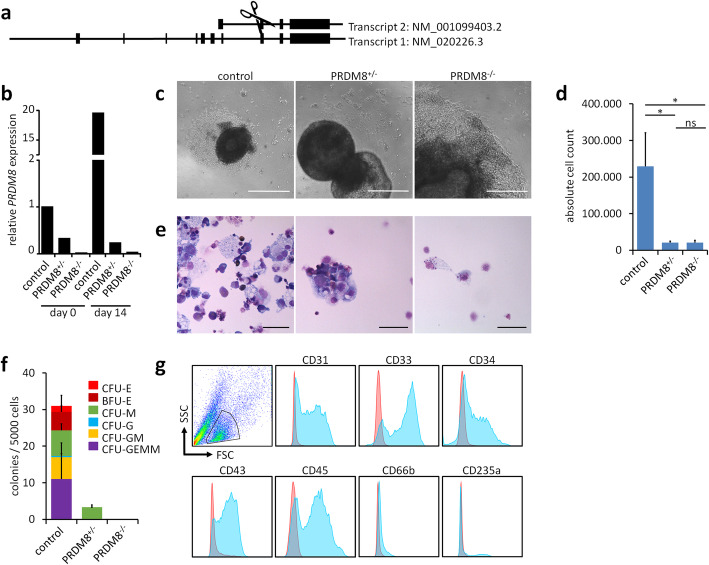
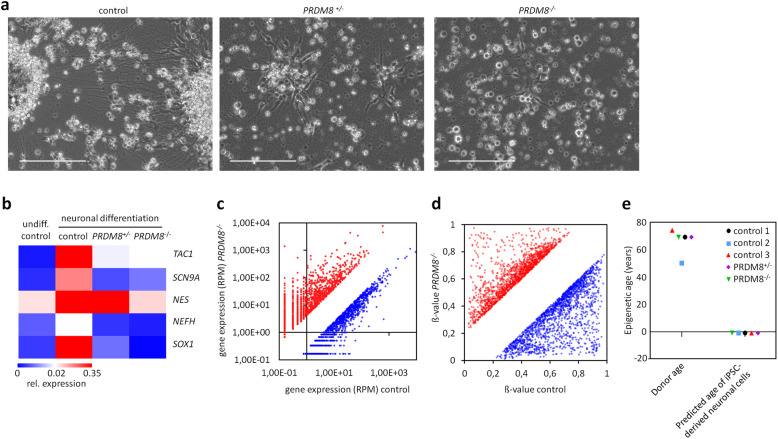
Results: In this study, we analyzed blood samples of 70 AA and 18 DKC patients to demonstrate that their epigenetic age predictions are overall increased, albeit not directly correlated with telomere length. Aberrant DNA methylation was observed in the gene PRDM8 in DKC and AA as well as in other diseases with premature aging phenotype, such as Down syndrome and Hutchinson-Gilford-Progeria syndrome. Aberrant DNA methylation patterns were particularly found within subsets of cell populations in DKC and AA samples as measured with barcoded bisulfite amplicon sequencing (BBA-seq). To gain insight into the functional relevance of PRDM8, we used CRISPR/Cas9 technology to generate induced pluripotent stem cells (iPSCs) with heterozygous and homozygous knockout. Loss of PRDM8 impaired hematopoietic and neuronal differentiation of iPSCs, even in the heterozygous knockout clone, but it did not impact on epigenetic age.
Conclusion: Taken together, our results demonstrate that epigenetic aging is accelerated in DKC and AA, independent from telomere attrition. Furthermore, aberrant DNA methylation in PRDM8 provides another biomarker for bone marrow failure syndromes and modulation of this gene in cellular subsets may be related to the hematopoietic and neuronal phenotypes observed in premature aging syndromes.
Keywords: Aging; Aplastic anemia; DNA methylation; Dyskeratosis congenita; Epigenetic clock; Hematopoietic differentiation; Neuronal differentiation; PRDM8; Telomere; iPSC.
Conflict of interest statement
WW and JF are involved in Cygenia GmbH (
Figures
Similar articles
-
DNA methylation in PRDM8 is indicative for dyskeratosis congenita.Oncotarget. 2016 Mar 8;7(10):10765-72. doi: 10.18632/oncotarget.7458. Oncotarget. 2016. PMID: 26909595 Free PMC article.
-
p53 Mediates Failure of Human Definitive Hematopoiesis in Dyskeratosis Congenita.Stem Cell Reports. 2017 Aug 8;9(2):409-418. doi: 10.1016/j.stemcr.2017.06.015. Epub 2017 Jul 27. Stem Cell Reports. 2017. PMID: 28757166 Free PMC article.
-
Aging of blood can be tracked by DNA methylation changes at just three CpG sites.Genome Biol. 2014 Feb 3;15(2):R24. doi: 10.1186/gb-2014-15-2-r24. Genome Biol. 2014. PMID: 24490752 Free PMC article.
-
Evaluation and Management of Hematopoietic Failure in Dyskeratosis Congenita.Hematol Oncol Clin North Am. 2018 Aug;32(4):669-685. doi: 10.1016/j.hoc.2018.04.003. Epub 2018 May 28. Hematol Oncol Clin North Am. 2018. PMID: 30047419 Free PMC article. Review.
-
Telomeres and marrow failure.Hematology Am Soc Hematol Educ Program. 2009:338-43. doi: 10.1182/asheducation-2009.1.338. Hematology Am Soc Hematol Educ Program. 2009. PMID: 20008219 Review.
Cited by
-
FAM111A Is a Novel Molecular Marker for Oocyte Aging.Biomedicines. 2022 Jan 25;10(2):257. doi: 10.3390/biomedicines10020257. Biomedicines. 2022. PMID: 35203468 Free PMC article.
-
Therapy Development by Genome Editing of Hematopoietic Stem Cells.Cells. 2021 Jun 14;10(6):1492. doi: 10.3390/cells10061492. Cells. 2021. PMID: 34198536 Free PMC article. Review.
-
The duality of PRDM proteins: epigenetic and structural perspectives.FEBS J. 2022 Mar;289(5):1256-1275. doi: 10.1111/febs.15844. Epub 2021 May 19. FEBS J. 2022. PMID: 33774927 Free PMC article. Review.
-
Transformative Materials to Create 3D Functional Human Tissue Models In Vitro in a Reproducible Manner.Adv Healthc Mater. 2023 Aug;12(20):e2301030. doi: 10.1002/adhm.202301030. Epub 2023 Jun 13. Adv Healthc Mater. 2023. PMID: 37311209 Free PMC article.
-
Recent advances in understanding telomere diseases.Fac Rev. 2022 Oct 19;11:31. doi: 10.12703/r/11-31. eCollection 2022. Fac Rev. 2022. PMID: 36311538 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
