

Loss of the seipin gene perturbs eggshell formation in Caenorhabditiselegans
- PMID: 32820022
- PMCID: PMC7578359
- DOI: 10.1242/dev.192997
Loss of the seipin gene perturbs eggshell formation in Caenorhabditiselegans
Abstract
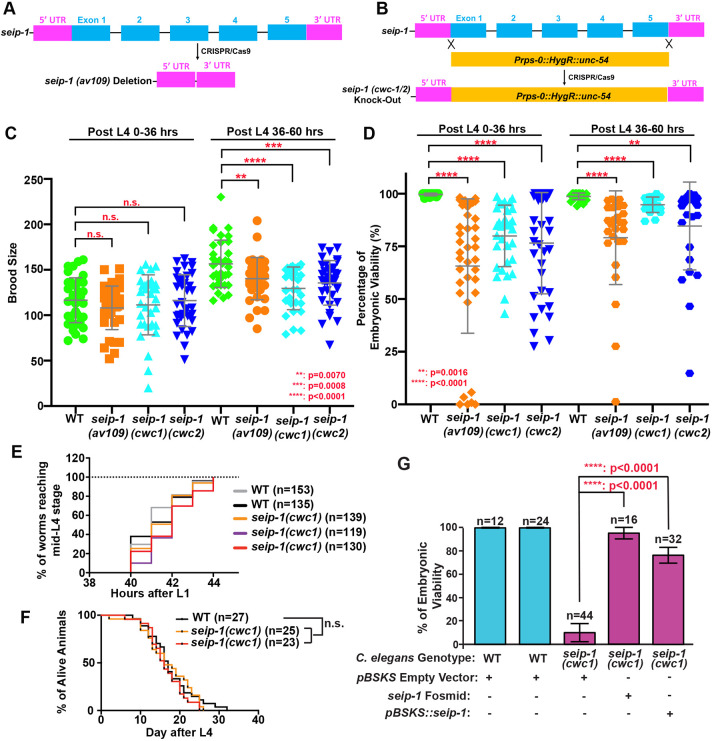
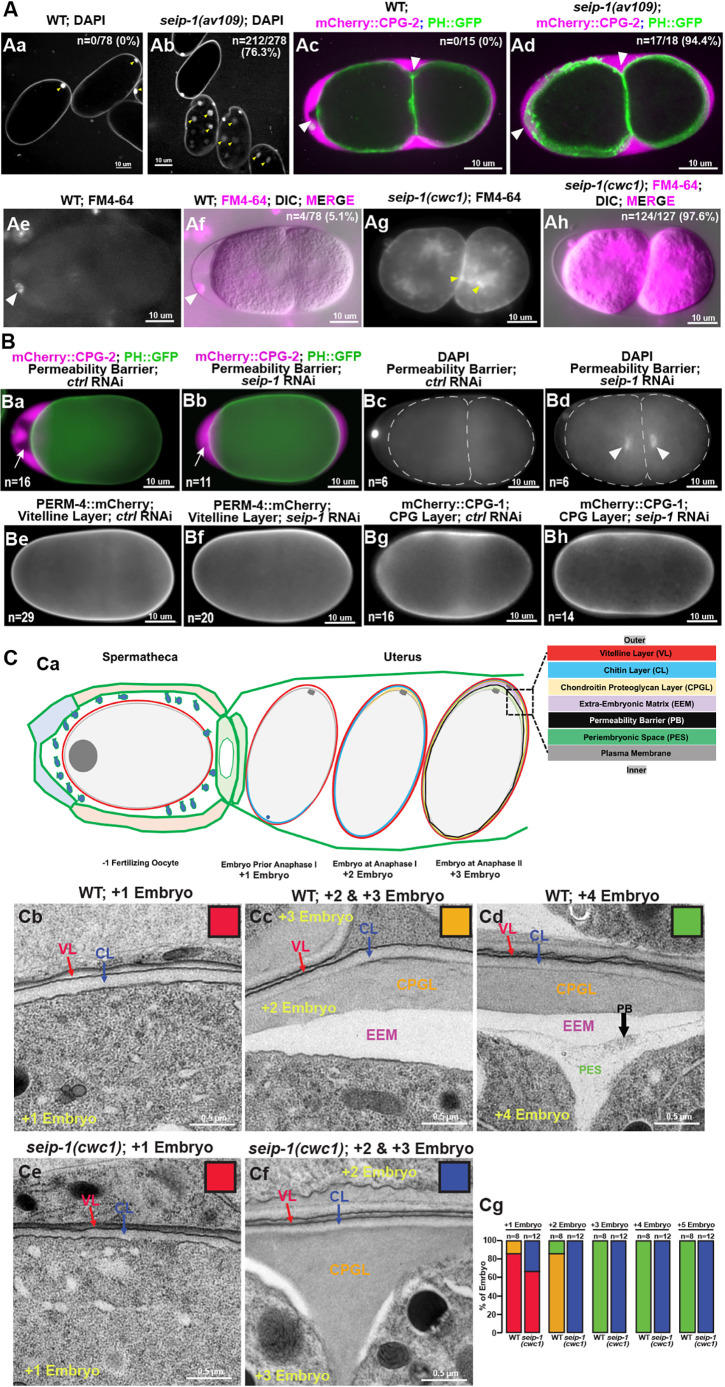
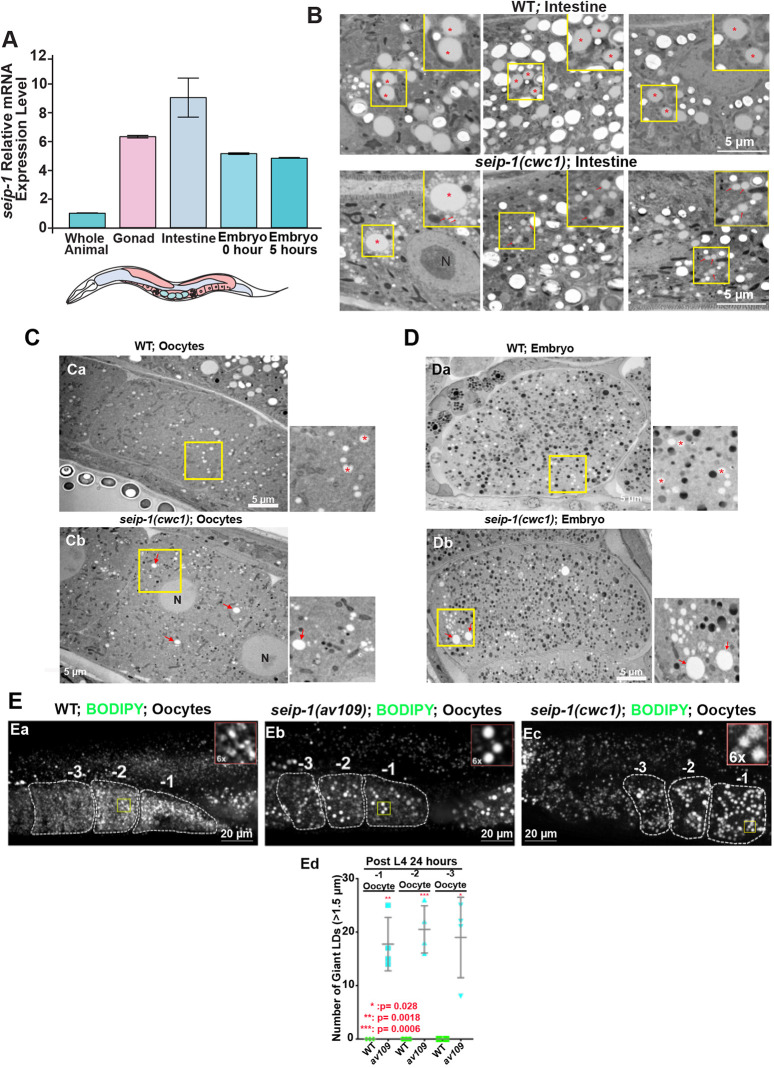
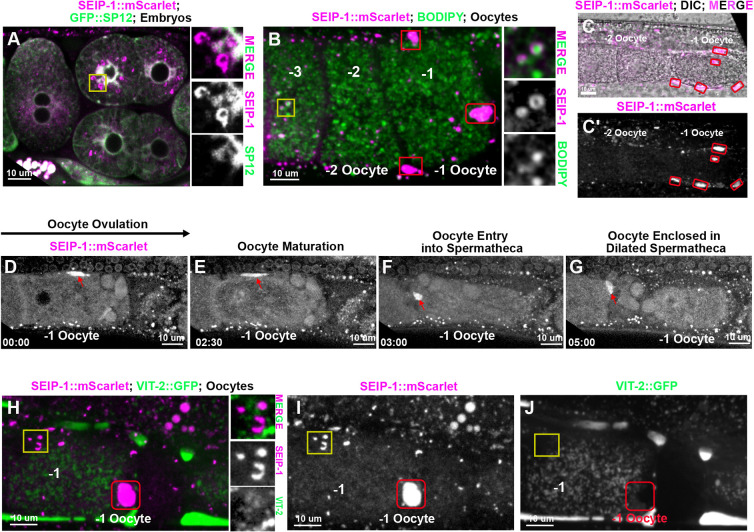
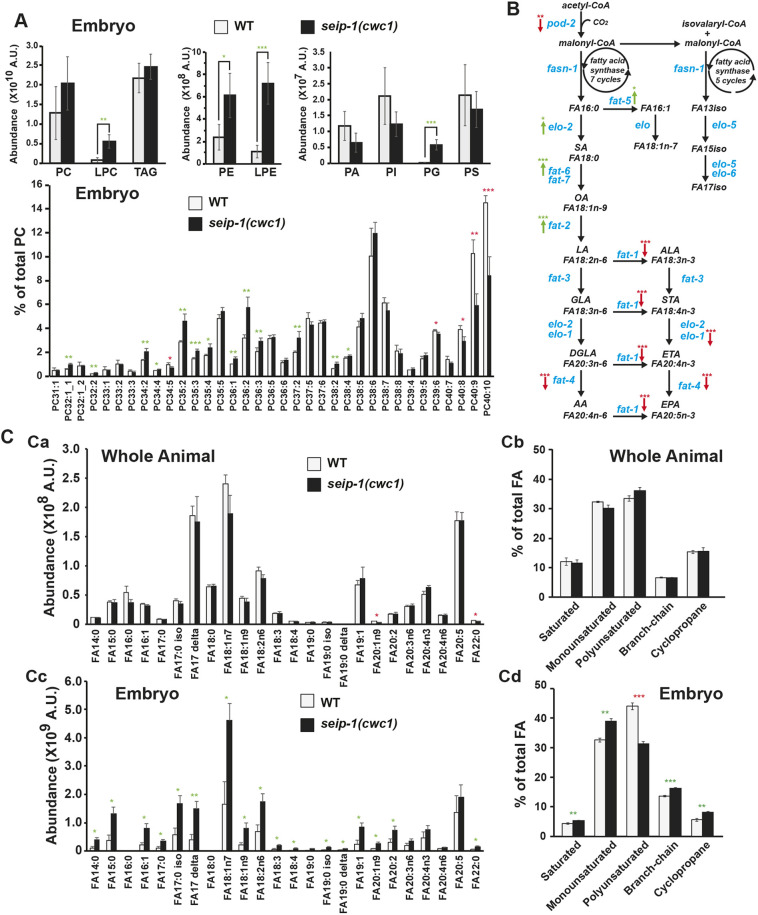
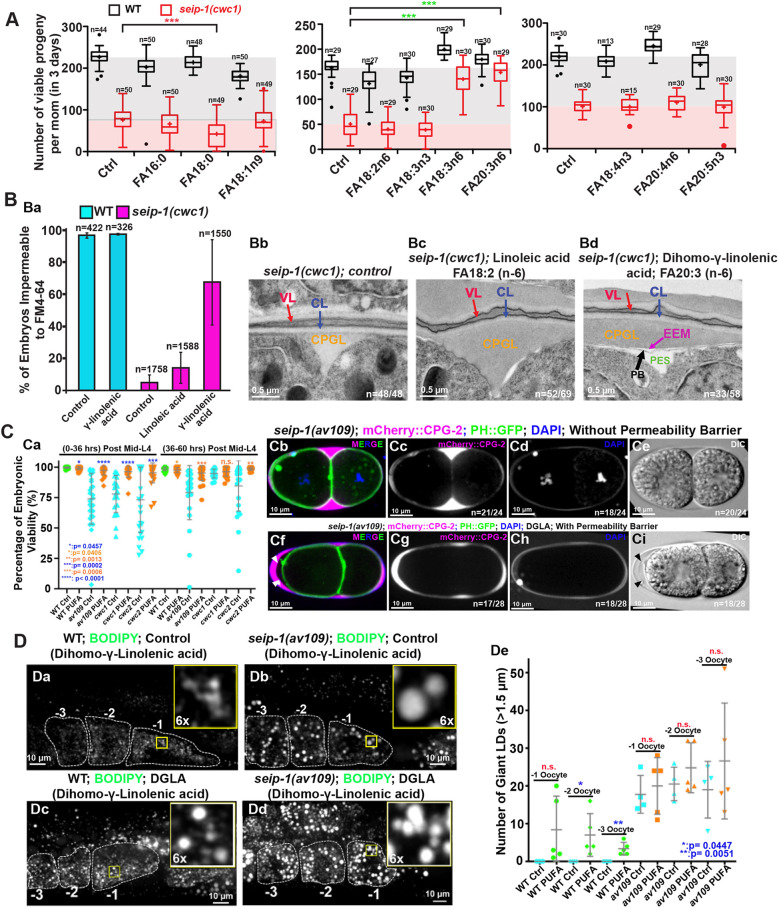
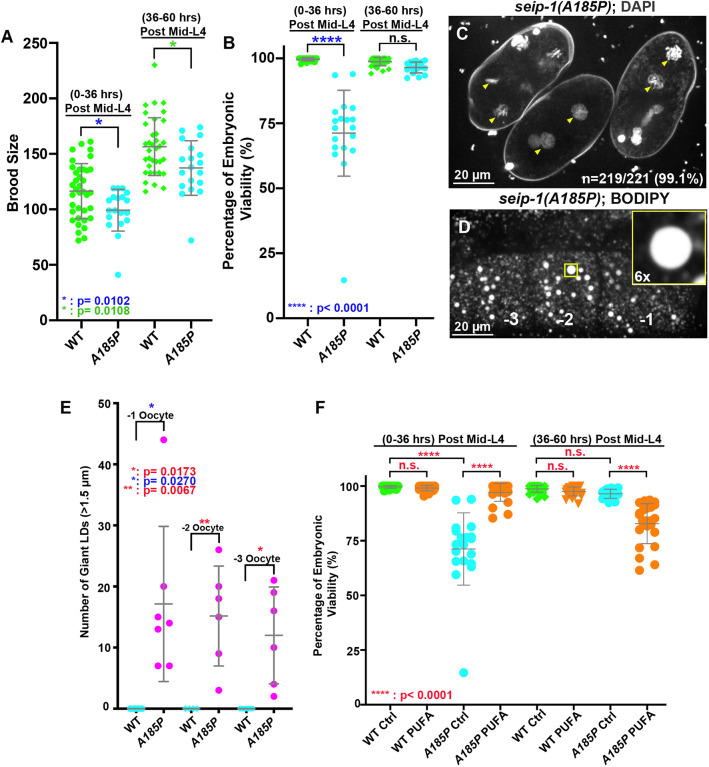
Seipin, an evolutionary conserved protein, plays pivotal roles during lipid droplet (LD) biogenesis and is associated with various human diseases with unclear mechanisms. Here, we analyzed Caenorhabditis elegans mutants deleted of the sole SEIPIN gene, seip-1 Homozygous seip-1 mutants displayed penetrant embryonic lethality, which is caused by the disruption of the lipid-rich permeability barrier, the innermost layer of the C. elegans embryonic eggshell. In C. elegans oocytes and embryos, SEIP-1 is associated with LDs and is crucial for controlling LD size and lipid homeostasis. The seip-1 deletion mutants reduced the ratio of polyunsaturated fatty acids (PUFAs) in their embryonic fatty acid pool. Interestingly, dietary supplementation of selected n-6 PUFAs rescued the embryonic lethality and defective permeability barrier. Accordingly, we propose that SEIP-1 may maternally regulate LD biogenesis and lipid homeostasis to orchestrate the formation of the permeability barrier for eggshell synthesis during embryogenesis. A lipodystrophy allele of seip-1 resulted in embryonic lethality as well and could be rescued by PUFA supplementation. These experiments support a great potential for using C. elegans to model SEIPIN-associated human diseases.
Keywords: Eggshell; Fatty acid; Lipid droplet; PUFAs; Permeability barrier; Seipin.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Benenati G., Penkov S., Müller-Reichert T., Entchev E. V. and Kurzchalia T. V. (2009). Two cytochrome P450s in Caenorhabditis elegans are essential for the organization of eggshell, correct execution of meiosis and the polarization of embryo. Mech. Dev. 126, 382-393. 10.1016/j.mod.2009.02.001 - DOI - PubMed
-
- Cutler R. G., Thompson K. W., Camandola S., Mack K. T. and Mattson M. P. (2014). Sphingolipid metabolism regulates development and lifespan in Caenorhabditis elegans. Mech. Ageing Dev. 143-144, 9-18. 10.1016/j.mad.2014.11.002 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
