

Phosphodiester modifications in mRNA poly(A) tail prevent deadenylation without compromising protein expression
- PMID: 32820035
- PMCID: PMC7668260
- DOI: 10.1261/rna.077099.120
Phosphodiester modifications in mRNA poly(A) tail prevent deadenylation without compromising protein expression
Abstract
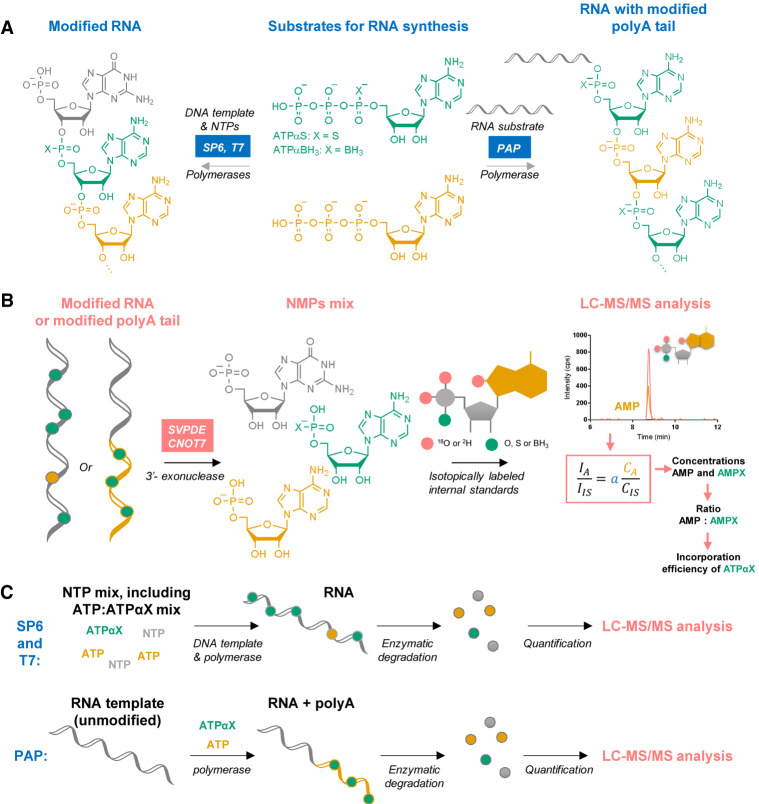
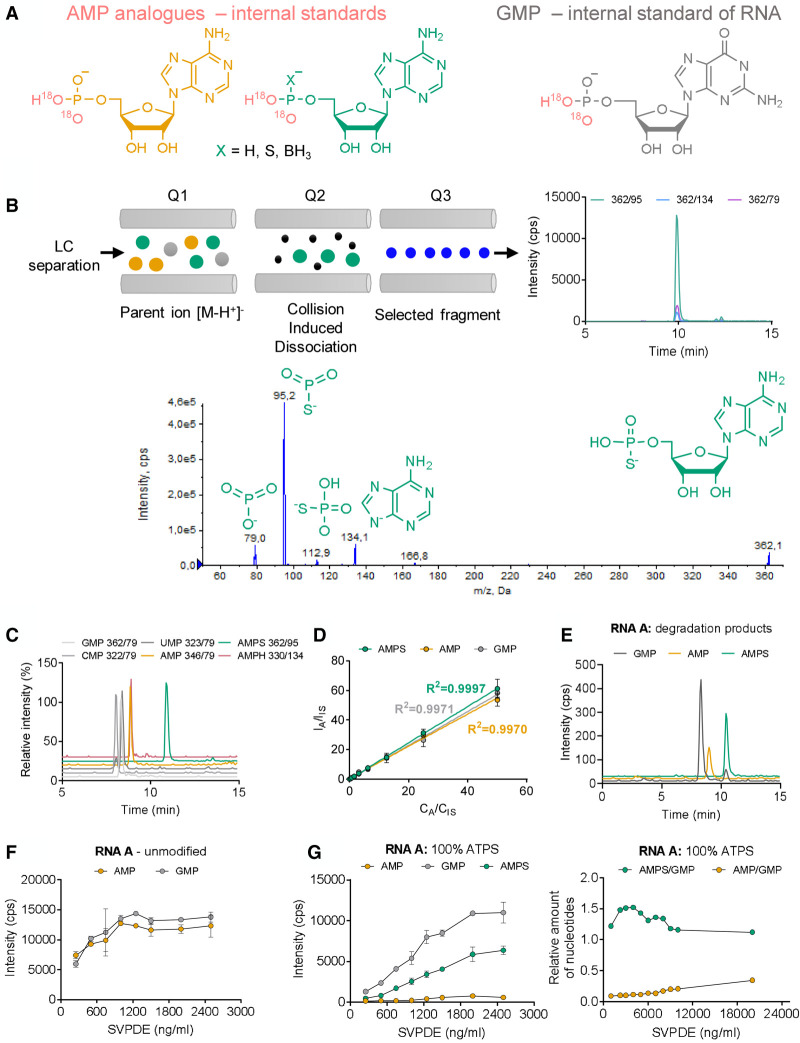
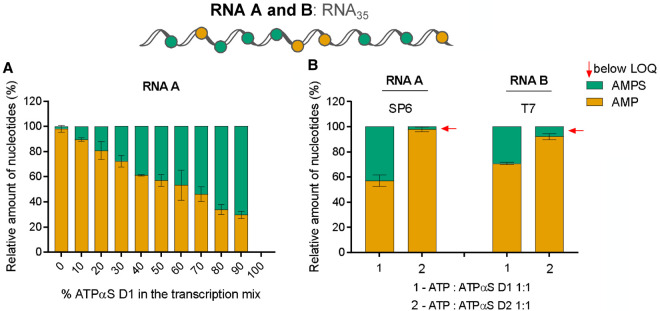
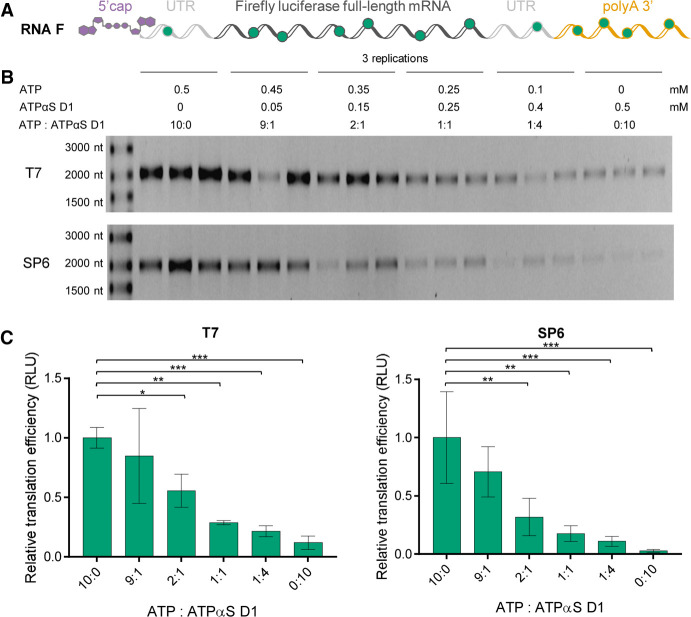
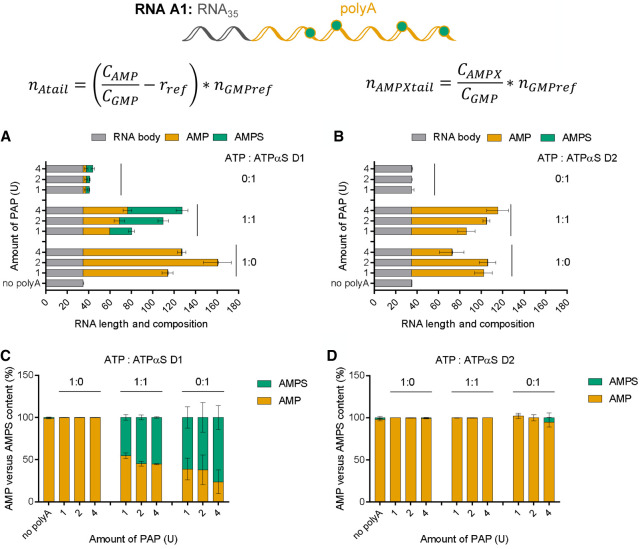
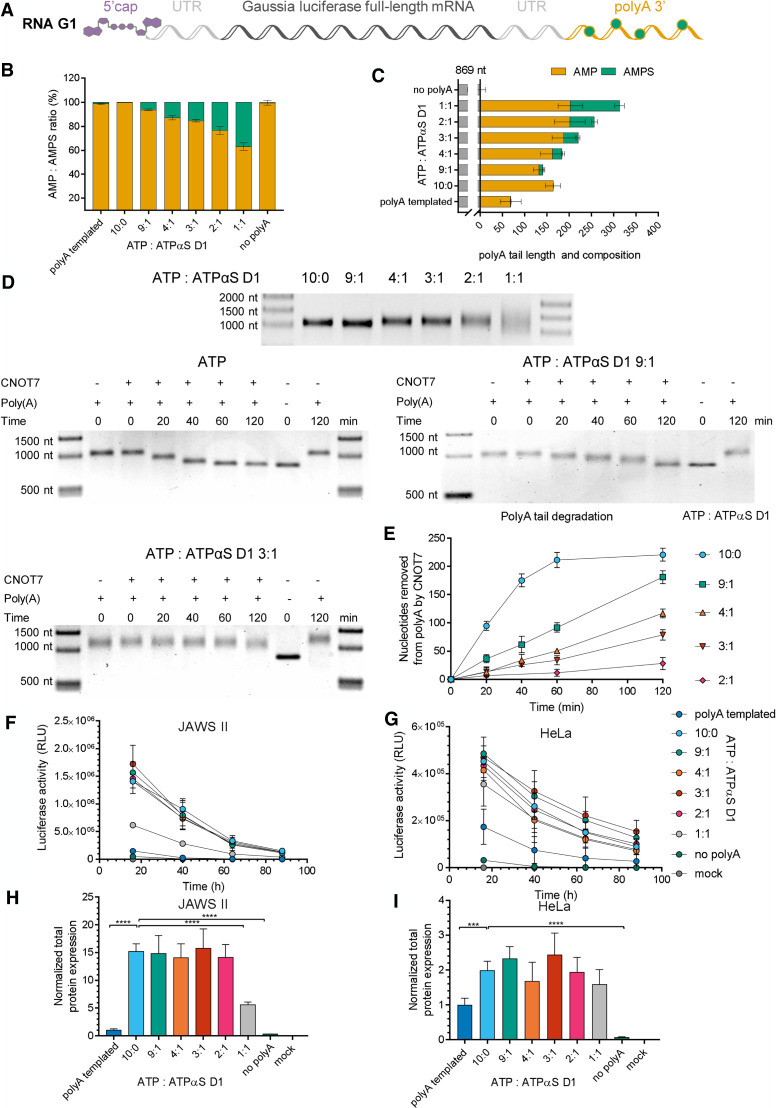
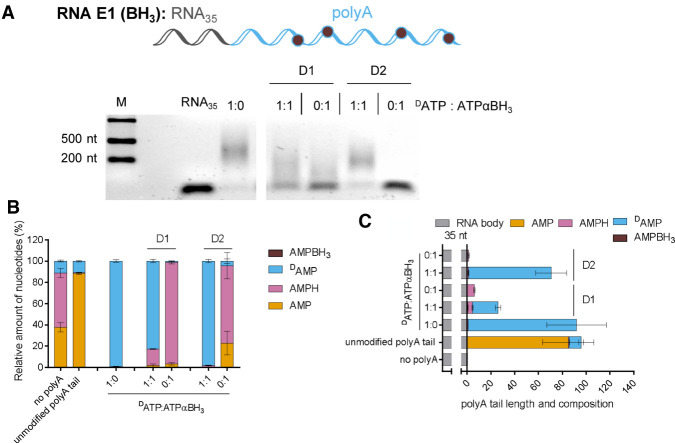
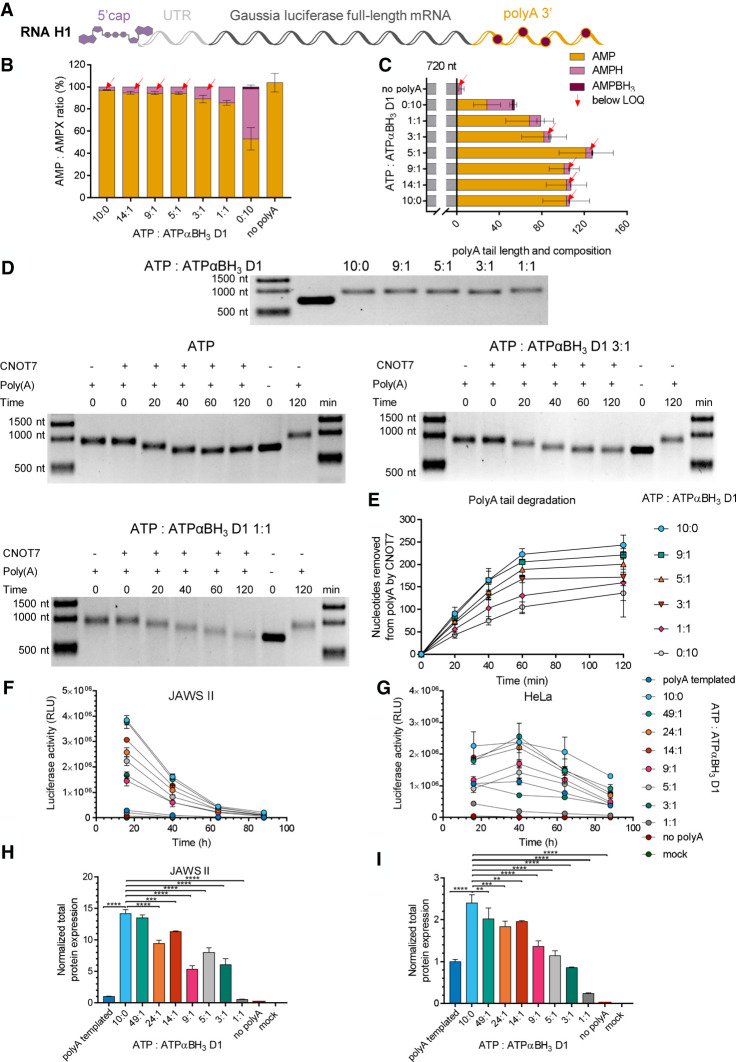
Chemical modifications enable preparation of mRNAs with augmented stability and translational activity. In this study, we explored how chemical modifications of 5',3'-phosphodiester bonds in the mRNA body and poly(A) tail influence the biological properties of eukaryotic mRNA. To obtain modified and unmodified in vitro transcribed mRNAs, we used ATP and ATP analogs modified at the α-phosphate (containing either O-to-S or O-to-BH3 substitutions) and three different RNA polymerases-SP6, T7, and poly(A) polymerase. To verify the efficiency of incorporation of ATP analogs in the presence of ATP, we developed a liquid chromatography-tandem mass spectrometry (LC-MS/MS) method for quantitative assessment of modification frequency based on exhaustive degradation of the transcripts to 5'-mononucleotides. The method also estimated the average poly(A) tail lengths, thereby providing a versatile tool for establishing a structure-biological property relationship for mRNA. We found that mRNAs containing phosphorothioate groups within the poly(A) tail were substantially less susceptible to degradation by 3'-deadenylase than unmodified mRNA and were efficiently expressed in cultured cells, which makes them useful research tools and potential candidates for future development of mRNA-based therapeutics.
Keywords: deadenylation; mRNA modification; resistance; transcription; translation.
© 2020 Strzelecka et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources