

KSHV LANA acetylation-selective acidic domain reader sequence mediates virus persistence
- PMID: 32820070
- PMCID: PMC7486799
- DOI: 10.1073/pnas.2004809117
KSHV LANA acetylation-selective acidic domain reader sequence mediates virus persistence
Abstract
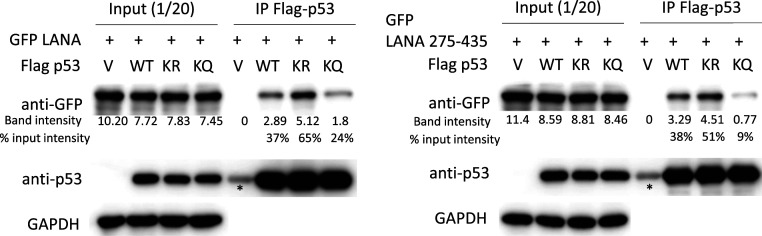
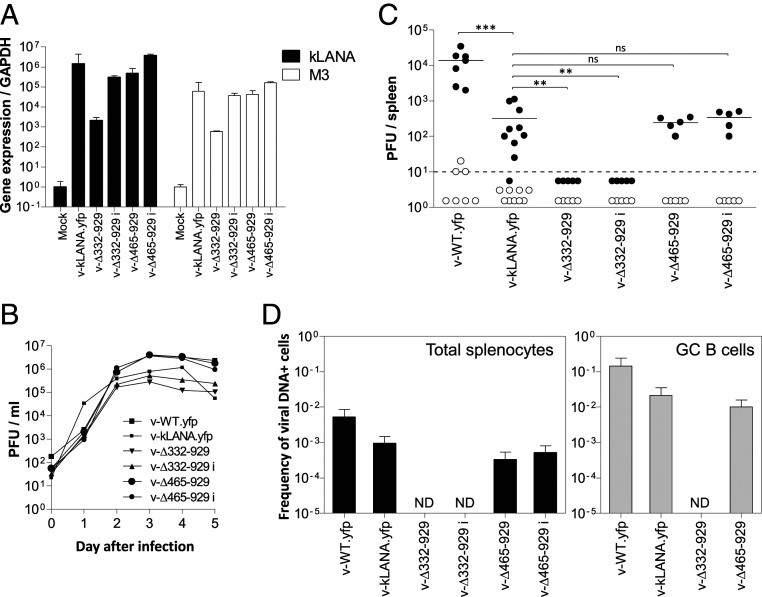
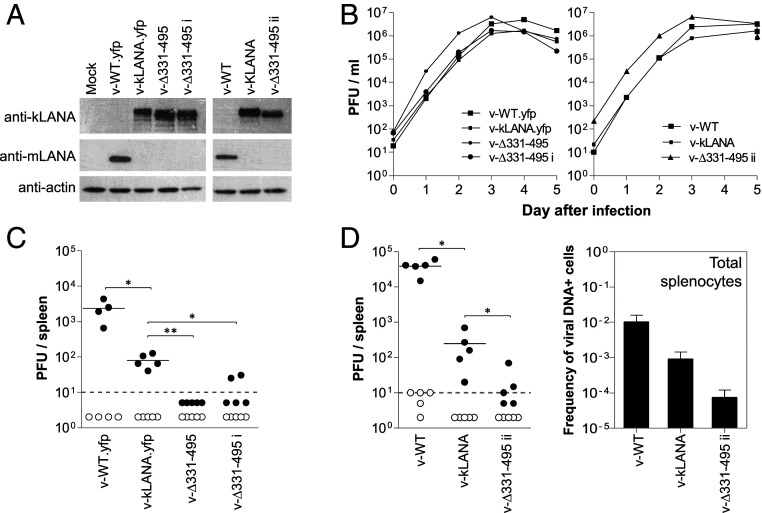
Viruses modulate biochemical cellular pathways to permit infection. A recently described mechanism mediates selective protein interactions between acidic domain readers and unacetylated, lysine-rich regions, opposite of bromodomain function. Kaposi´s sarcoma (KS)-associated herpesvirus (KSHV) is tightly linked with KS, primary effusion lymphoma, and multicentric Castleman's disease. KSHV latently infects cells, and its genome persists as a multicopy, extrachromosomal episome. During latency, KSHV expresses a small subset of genes, including the latency-associated nuclear antigen (LANA), which mediates viral episome persistence. Here we show that LANA contains two tandem, partially overlapping, acidic domain sequences homologous to the SET oncoprotein acidic domain reader. This domain selectively interacts with unacetylated p53, as evidenced by reduced LANA interaction after overexpression of CBP, which acetylates p53, or with an acetylation mimicking carboxyl-terminal domain p53 mutant. Conversely, the interaction of LANA with an acetylation-deficient p53 mutant is enhanced. Significantly, KSHV LANA mutants lacking the acidic domain reader sequence are deficient for establishment of latency and persistent infection. This deficiency was confirmed under physiological conditions, on infection of mice with a murine gammaherpesvirus 68 chimera expressing LANA, where the virus was highly deficient in establishing latent infection in germinal center B cells. Therefore, LANA's acidic domain reader is critical for viral latency. These results implicate an acetylation-dependent mechanism mediating KSHV persistence and expand the role of acidic domain readers.
Keywords: Kaposi´s sarcoma herpesvirus; acetylation-regulated interaction; acidic domain reader; latency-associated nuclear antigen; virus persistence.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
IFI16 recruits HDAC1 and HDAC2 to deacetylate the Kaposi's sarcoma-associated herpesvirus (KSHV) latency-associated nuclear antigen (LANA), facilitating latency.J Virol. 2025 Mar 18;99(3):e0154924. doi: 10.1128/jvi.01549-24. Epub 2025 Feb 10. J Virol. 2025. PMID: 39927772 Free PMC article.
-
Kaposi's Sarcoma-Associated Herpesvirus LANA-Adjacent Regions with Distinct Functions in Episome Segregation or Maintenance.J Virol. 2019 Mar 5;93(6):e02158-18. doi: 10.1128/JVI.02158-18. Print 2019 Mar 15. J Virol. 2019. PMID: 30626680 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus-encoded LANA contributes to viral latent replication by activating phosphorylation of survivin.J Virol. 2014 Apr;88(8):4204-17. doi: 10.1128/JVI.03855-13. Epub 2014 Jan 29. J Virol. 2014. PMID: 24478433 Free PMC article.
-
The latency-associated nuclear antigen, a multifunctional protein central to Kaposi's sarcoma-associated herpesvirus latency.Future Microbiol. 2011 Dec;6(12):1399-413. doi: 10.2217/fmb.11.137. Future Microbiol. 2011. PMID: 22122438 Free PMC article. Review.
-
Kaposi's Sarcoma Herpesvirus Genome Persistence.Front Microbiol. 2016 Aug 12;7:1149. doi: 10.3389/fmicb.2016.01149. eCollection 2016. Front Microbiol. 2016. PMID: 27570517 Free PMC article. Review.
Cited by
-
Deciphering the acetylation code of p53 in transcription regulation and tumor suppression.Oncogene. 2022 May;41(22):3039-3050. doi: 10.1038/s41388-022-02331-9. Epub 2022 Apr 29. Oncogene. 2022. PMID: 35487975 Free PMC article. Review.
-
Endoplasmic Reticulum-Shaping Atlastin Proteins Facilitate KSHV Replication.Front Cell Infect Microbiol. 2022 Jan 13;11:790243. doi: 10.3389/fcimb.2021.790243. eCollection 2021. Front Cell Infect Microbiol. 2022. PMID: 35096644 Free PMC article.
-
Vital for Viruses: Intrinsically Disordered Proteins.J Mol Biol. 2023 Jun 1;435(11):167860. doi: 10.1016/j.jmb.2022.167860. Epub 2023 Jun 16. J Mol Biol. 2023. PMID: 37330280 Free PMC article. Review.
-
RNA-guided gene editing of the murine gammaherpesvirus 68 genome reduces infectious virus production.PLoS One. 2021 Jun 4;16(6):e0252313. doi: 10.1371/journal.pone.0252313. eCollection 2021. PLoS One. 2021. PMID: 34086743 Free PMC article.
-
Sirtuin 2 promotes human cytomegalovirus replication by regulating cell cycle progression.mSystems. 2023 Dec 21;8(6):e0051023. doi: 10.1128/msystems.00510-23. Epub 2023 Nov 2. mSystems. 2023. PMID: 37916830 Free PMC article.
References
-
- Chang Y. et al. ., Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi’s sarcoma. Science 266, 1865–1869 (1994). - PubMed
-
- Cesarman E., Chang Y., Moore P. S., Said J. W., Knowles D. M., Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N. Engl. J. Med. 332, 1186–1191 (1995). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
