

Autophagy regulates trophoblast invasion by targeting NF-κB activity
- PMID: 32820194
- PMCID: PMC7441061
- DOI: 10.1038/s41598-020-70959-2
Autophagy regulates trophoblast invasion by targeting NF-κB activity
Abstract
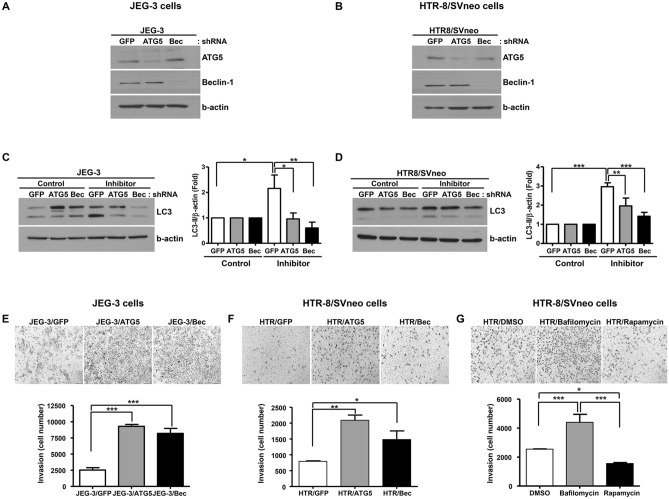
Preeclampsia is one of the most serious complications of pregnancy, affecting 5-10% of parturients worldwide. Recent studies have suggested that autophagy is involved in trophoblast invasion and may be associated with defective placentation underlying preeclampsia. We thus aimed to understand the mechanistic link between autophagy and trophoblast invasion. Using the two most commonly used trophoblast cell lines, JEG-3 and HTR-8/SVneo, we inhibited autophagy by ATG5 and beclin-1 shRNA. Conversion of LC3-II was evaluated in ATG5 and beclin-1 knock-down cells in the presence of the lysosomal protease inhibitors E-64d and pepstatin A, to detect the efficiency of autophagy inhibition. Upon autophagy inhibition, we measured cell invasion, activity of NF-κB and related signaling pathways, MMP-2, MMP-9, sFlt-1, and TNF-α levels. Autophagy inhibition increased the invasiveness of these trophoblastic cell lines and increased Akt and NF-κB activity as well as p65 expression. Of note, an NF-κB inhibitor significantly attenuated the trophoblast invasion induced by autophagy inhibition. Autophagy inhibition was also associated with increased MMP-2 and MMP-9 levels and decreased the production of sFlt-1 and TNF-α. Collectively, our results indicate that autophagy regulates trophoblast invasiveness in which the NF-κB pathway and MMP-2, MMP-9, sFlt-1 and TNF-α levels are affected.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Kadyrov M, Kingdom JC, Huppertz B. Divergent trophoblast invasion and apoptosis in placental bed spiral arteries from pregnancies complicated by maternal anemia and early-onset preeclampsia/intrauterine growth restriction. Am. J. Obstet. Gynecol. 2006;194:557–563. doi: 10.1016/j.ajog.2005.07.035. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
