

Kinetochore-microtubule coupling mechanisms mediated by the Ska1 complex and Cdt1
- PMID: 32844209
- PMCID: PMC7591161
- DOI: 10.1042/EBC20190075
Kinetochore-microtubule coupling mechanisms mediated by the Ska1 complex and Cdt1
Abstract
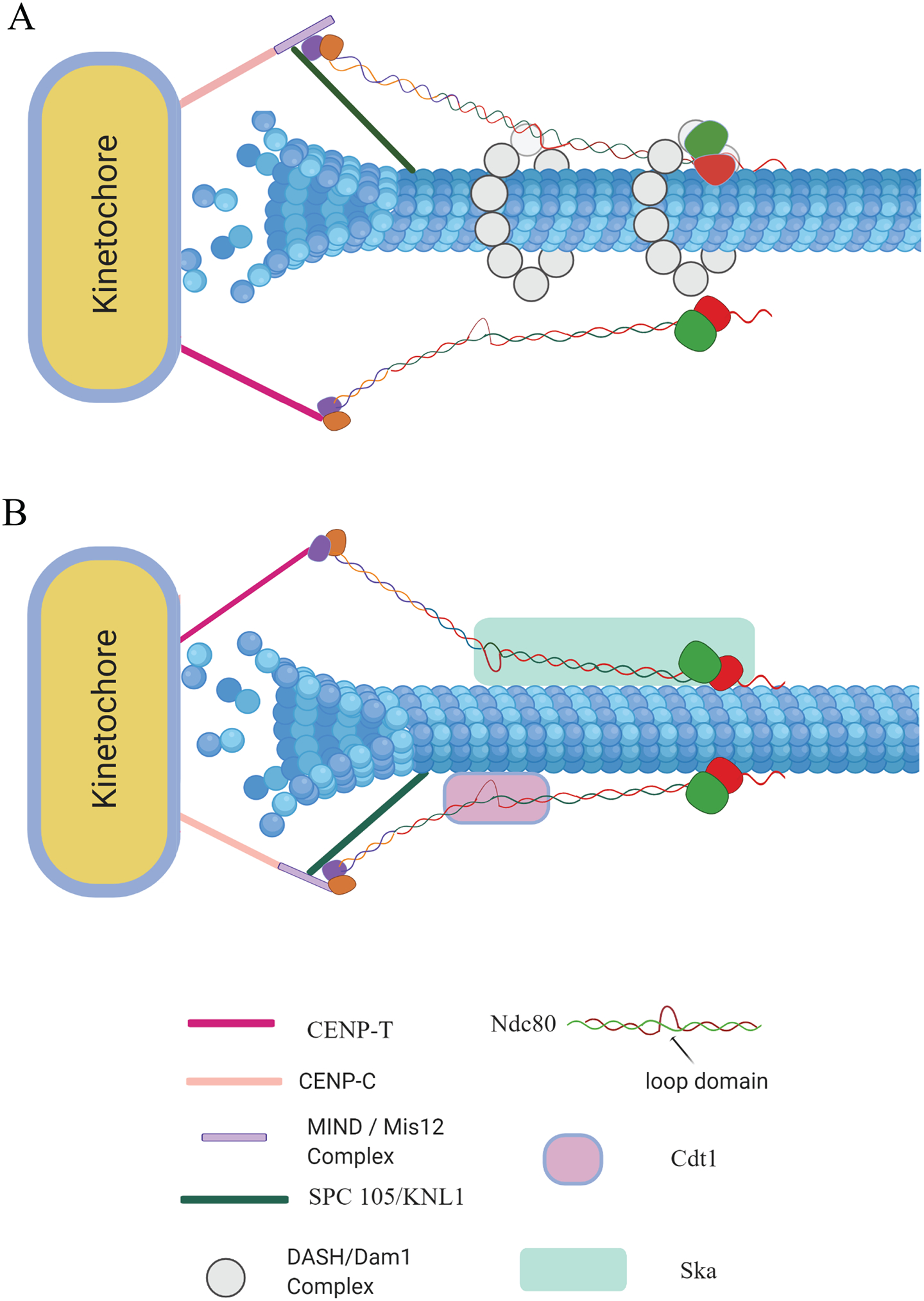
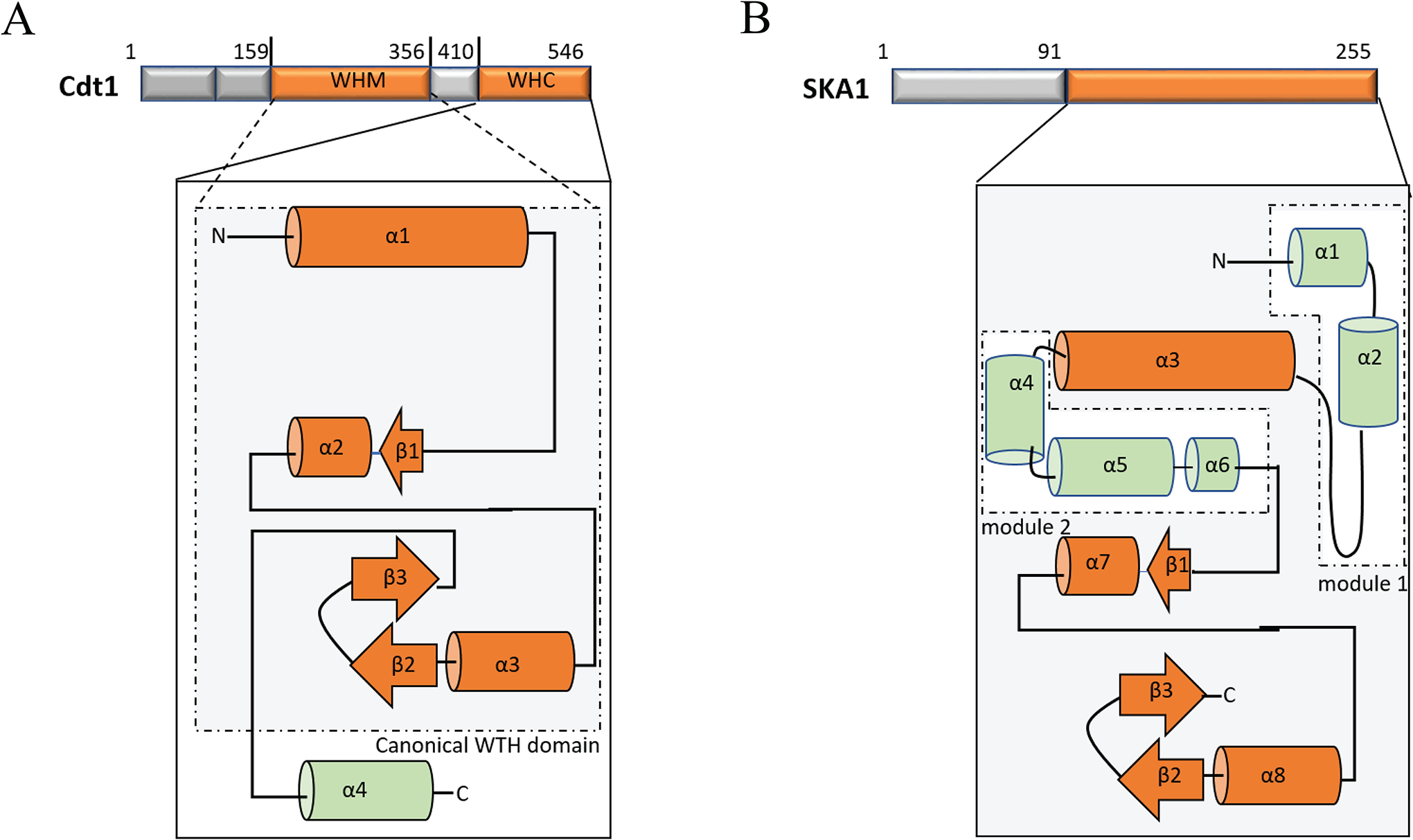
The faithful segregation of duplicated sister chromatids rely on the remarkable ability of kinetochores to sustain stable load bearing attachments with the dynamic plus ends of kinetochore-microtubules (kMTs). The outer layer of the kinetochore recruits several motor and non-motor microtubule-associated proteins (MAPs) that help the kinetochores establish and maintain a load bearing dynamic attachment with kMTs. The primary kMT-binding protein, the Ndc80 complex (Ndc80c), which is highly conserved among diverse organisms from yeast to humans, performs this essential function with assistance from other MAPs. These MAPs are not an integral part of the kinetochore, but they localize to the kinetochore periodically throughout mitosis and regulate the strength of the kinetochore microtubule attachments. Here, we attempt to summarize the recent advances that have been made toward furthering our understanding of this co-operation between the Ndc80c and these MAPs, focusing on the spindle and kinetochore-associated 1 (Ska1) complex (Ska1c) and Cdc10-dependent transcript 1 (Cdt1) in humans.
Keywords: Chromosomes; Kinetochores; MAPs; Microtubules; Mitosis; Ndc80.
© 2020 The Author(s). Published by Portland Press Limited on behalf of the Biochemical Society.
Conflict of interest statement
Conflict of Interest
The authors declare no conflicts of interest.
Figures
Similar articles
-
The Ndc80-Cdt1-Ska1 complex is a central processive kinetochore-microtubule coupling unit.J Cell Biol. 2023 Aug 7;222(8):e202208018. doi: 10.1083/jcb.202208018. Epub 2023 Jun 2. J Cell Biol. 2023. PMID: 37265445 Free PMC article.
-
Ska3 Phosphorylated by Cdk1 Binds Ndc80 and Recruits Ska to Kinetochores to Promote Mitotic Progression.Curr Biol. 2017 May 22;27(10):1477-1484.e4. doi: 10.1016/j.cub.2017.03.060. Epub 2017 May 4. Curr Biol. 2017. PMID: 28479321
-
The Hec1/Ndc80 tail domain is required for force generation at kinetochores, but is dispensable for kinetochore-microtubule attachment formation and Ska complex recruitment.Mol Biol Cell. 2020 Jul 1;31(14):1453-1473. doi: 10.1091/mbc.E20-05-0286. Epub 2020 May 13. Mol Biol Cell. 2020. PMID: 32401635 Free PMC article.
-
Mapping the kinetochore MAP functions required for stabilizing microtubule attachments to chromosomes during metaphase.Cytoskeleton (Hoboken). 2019 Jun;76(6):398-412. doi: 10.1002/cm.21559. Epub 2019 Sep 9. Cytoskeleton (Hoboken). 2019. PMID: 31454167 Free PMC article. Review.
-
How the SAC gets the axe: Integrating kinetochore microtubule attachments with spindle assembly checkpoint signaling.Bioarchitecture. 2015;5(1-2):1-12. doi: 10.1080/19490992.2015.1090669. Epub 2015 Oct 2. Bioarchitecture. 2015. PMID: 26430805 Free PMC article. Review.
Cited by
-
SEC-SAXS/MC Ensemble Structural Studies of the Microtubule Binding Protein Cdt1 Show Monomeric, Folded-Over Conformations.Cytoskeleton (Hoboken). 2025 Jun;82(6):372-387. doi: 10.1002/cm.21954. Epub 2024 Nov 6. Cytoskeleton (Hoboken). 2025. PMID: 39503309 Free PMC article.
-
ETV5 overexpression promotes progression of esophageal squamous cell carcinoma by upregulating SKA1 and TRPV2.Int J Med Sci. 2022 Jun 21;19(6):1072-1081. doi: 10.7150/ijms.71892. eCollection 2022. Int J Med Sci. 2022. PMID: 35813298 Free PMC article.
-
The Ndc80-Cdt1-Ska1 complex is a central processive kinetochore-microtubule coupling unit.J Cell Biol. 2023 Aug 7;222(8):e202208018. doi: 10.1083/jcb.202208018. Epub 2023 Jun 2. J Cell Biol. 2023. PMID: 37265445 Free PMC article.
-
SAXS/MC studies of the mixed-folded protein Cdt1 reveal monomeric, folded over conformations.bioRxiv [Preprint]. 2024 Jan 3:2024.01.03.573975. doi: 10.1101/2024.01.03.573975. bioRxiv. 2024. Update in: Cytoskeleton (Hoboken). 2025 Jun;82(6):372-387. doi: 10.1002/cm.21954. PMID: 38260441 Free PMC article. Updated. Preprint.
References
-
- Cheeseman IM, Desai A. Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008;9(1):33–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources