

Unique Structural Features of Mammalian NEIL2 DNA Glycosylase Prime Its Activity for Diverse DNA Substrates and Environments
- PMID: 32846144
- PMCID: PMC7796977
- DOI: 10.1016/j.str.2020.08.001
Unique Structural Features of Mammalian NEIL2 DNA Glycosylase Prime Its Activity for Diverse DNA Substrates and Environments
Abstract
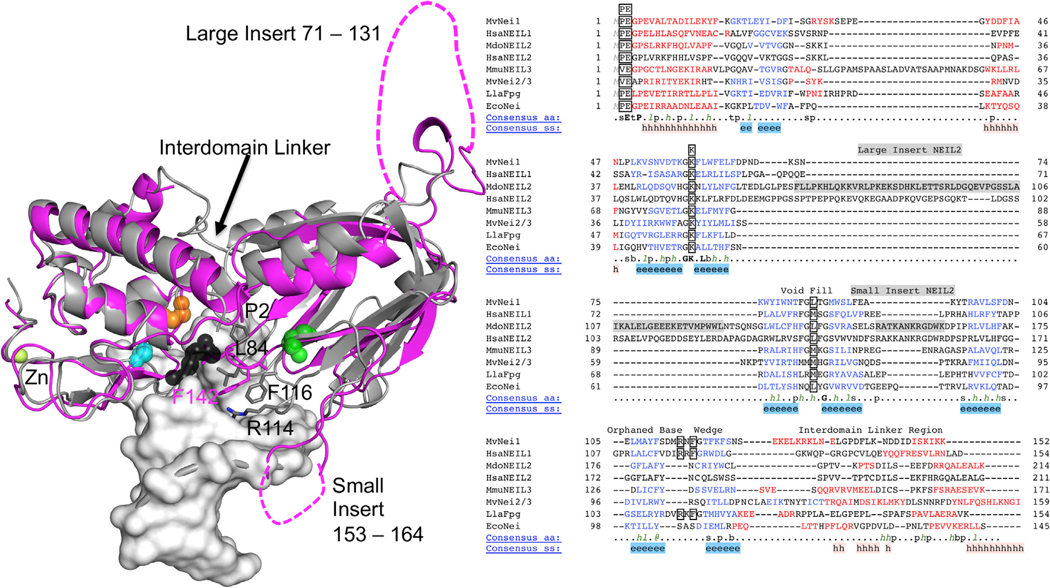
Oxidative damage on DNA arising from both endogenous and exogenous sources can result in base modifications that promote errors in replication as well as generating sites of base loss (abasic sites) that present unique challenges to maintaining genomic integrity. These lesions are excised by DNA glycosylases in the first step of the base excision repair pathway. Here we present the first crystal structure of a NEIL2 glycosylase, an enzyme active on cytosine oxidation products and abasic sites. The structure reveals an unusual "open" conformation not seen in NEIL1 or NEIL3 orthologs. NEIL2 is predicted to adopt a "closed" conformation when bound to its substrate. Combined crystallographic and solution-scattering studies show the enzyme to be conformationally dynamic in a manner distinct among the NEIL glycosylases and provide insight into the unique substrate preference of this enzyme. In addition, we characterized three cancer variants of human NEIL2, namely S140N, G230W, and G303R.
Keywords: DNA damage; DNA repair; NEIL2 glycosylase; SAXS; X-ray crystallography.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures





Comment in
-
Breaking the Rules: Protein Sculpting in NEIL2 Regulation.Structure. 2021 Jan 7;29(1):1-2. doi: 10.1016/j.str.2020.12.011. Structure. 2021. PMID: 33417890 Free PMC article.
Similar articles
-
Requirements for DNA bubble structure for efficient cleavage by helix-two-turn-helix DNA glycosylases.Mutagenesis. 2020 Feb 13;35(1):119-128. doi: 10.1093/mutage/gez047. Mutagenesis. 2020. PMID: 31784740
-
NEIL1 and NEIL2 Are Recruited as Potential Backup for OGG1 upon OGG1 Depletion or Inhibition by TH5487.Int J Mol Sci. 2021 Apr 27;22(9):4542. doi: 10.3390/ijms22094542. Int J Mol Sci. 2021. PMID: 33925271 Free PMC article.
-
Structural investigation of a viral ortholog of human NEIL2/3 DNA glycosylases.DNA Repair (Amst). 2013 Dec;12(12):1062-71. doi: 10.1016/j.dnarep.2013.09.004. Epub 2013 Oct 10. DNA Repair (Amst). 2013. PMID: 24120312 Free PMC article.
-
Functional roles and cancer variants of the bifunctional glycosylase NEIL2.Environ Mol Mutagen. 2024 Apr;65 Suppl 1:40-56. doi: 10.1002/em.22555. Epub 2023 Jun 26. Environ Mol Mutagen. 2024. PMID: 37310399 Review.
-
DNA glycosylase NEIL2 functions in multiple cellular processes.Prog Biophys Mol Biol. 2021 Sep;164:72-80. doi: 10.1016/j.pbiomolbio.2021.03.003. Epub 2021 Mar 20. Prog Biophys Mol Biol. 2021. PMID: 33753087 Free PMC article. Review.
Cited by
-
Distinct Mechanisms of Target Search by Endonuclease VIII-like DNA Glycosylases.Cells. 2022 Oct 11;11(20):3192. doi: 10.3390/cells11203192. Cells. 2022. PMID: 36291061 Free PMC article.
-
Structural and Evolutionary Adaptations of Nei-Like DNA Glycosylases Proteins Involved in Base Excision Repair of Oxidative DNA Damage in Vertebrates.Oxid Med Cell Longev. 2022 Apr 4;2022:1144387. doi: 10.1155/2022/1144387. eCollection 2022. Oxid Med Cell Longev. 2022. PMID: 35419164 Free PMC article.
-
Noncatalytic Domains in DNA Glycosylases.Int J Mol Sci. 2022 Jun 30;23(13):7286. doi: 10.3390/ijms23137286. Int J Mol Sci. 2022. PMID: 35806289 Free PMC article. Review.
-
Molecular Evolution of the Activating Transcription Factors Shapes the Adaptive Cellular Responses to Oxidative Stress.Oxid Med Cell Longev. 2022 Jul 13;2022:2153996. doi: 10.1155/2022/2153996. eCollection 2022. Oxid Med Cell Longev. 2022. Retraction in: Oxid Med Cell Longev. 2024 Jan 9;2024:9875345. doi: 10.1155/2024/9875345. PMID: 35873797 Free PMC article. Retracted.
-
Research progress on the role of the NEIL family in cancer.Front Cell Dev Biol. 2025 Jul 21;13:1612329. doi: 10.3389/fcell.2025.1612329. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40761744 Free PMC article. Review.
References
-
- Alexeeva M, Guragain P, Tesfahun AN, Tomkuviene M, Arshad A, Gerasimaite R, Ruksenaite A, Urbanaviciute G, Bjørå s M, Laerdahl JK, et al. (2018). Excision of the doubly methylated base N(4),5-dimethylcytosine from DNA by Escherichia coli Nei and Fpg proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 373, 20170337. - PMC - PubMed
-
- Bandaru V, Blaisdell JO, and Wallace SS (2006). Oxidative DNA glycosylases: recipes from cloning to characterization. Methods Enzymol. 408, 15–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
