

Thermus thermophilus Argonaute Functions in the Completion of DNA Replication
- PMID: 32846159
- PMCID: PMC7502556
- DOI: 10.1016/j.cell.2020.07.036
Thermus thermophilus Argonaute Functions in the Completion of DNA Replication
Abstract
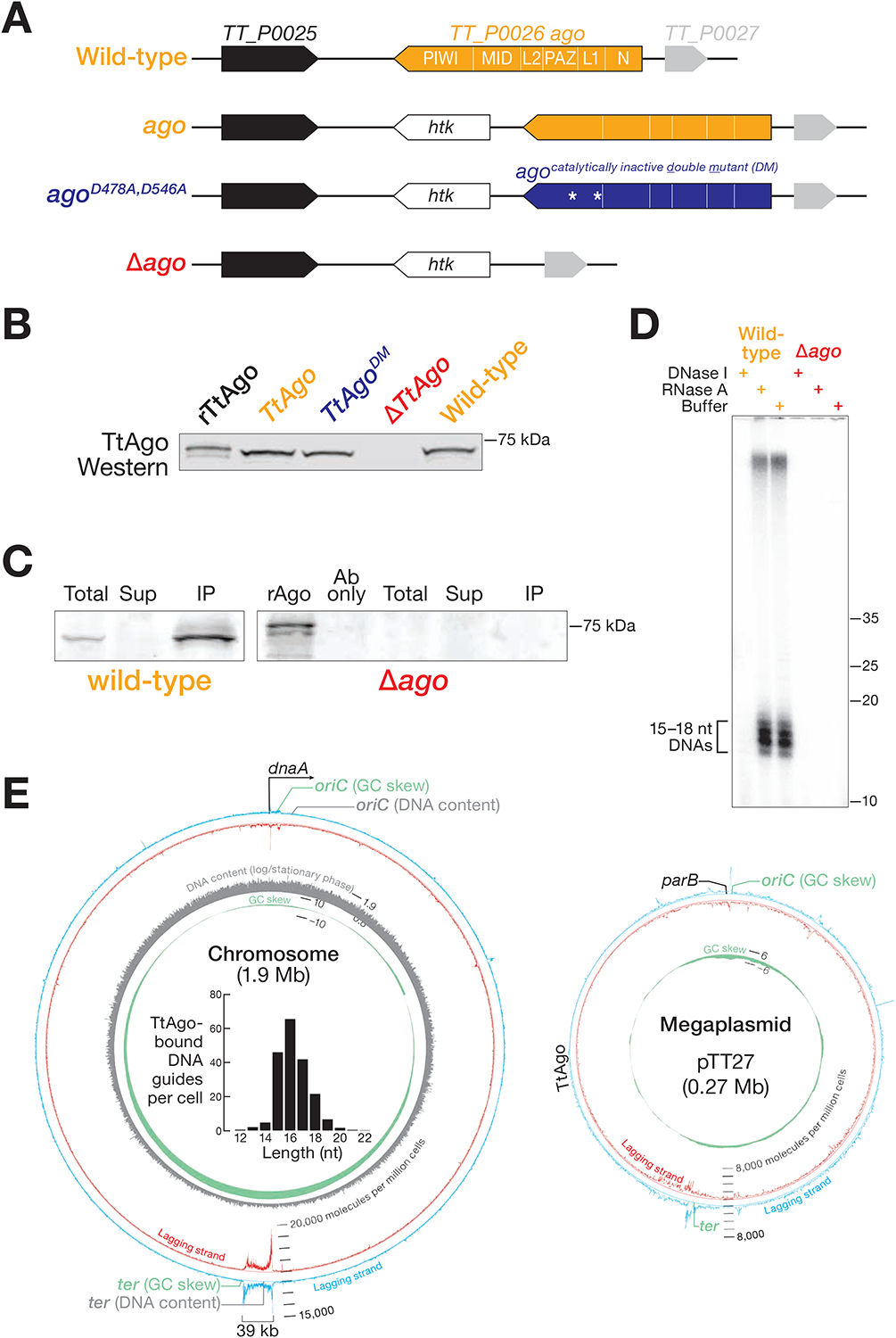
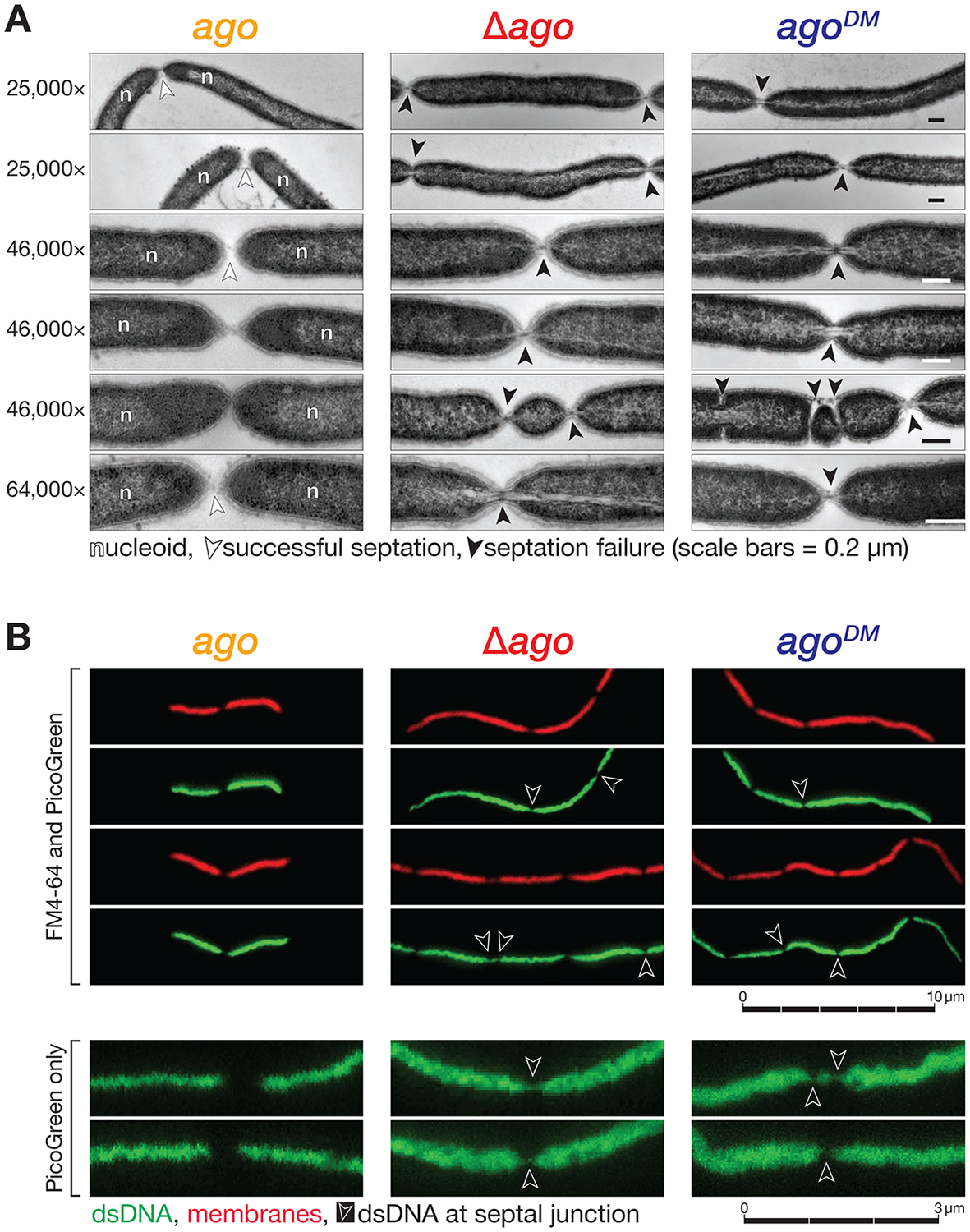
In many eukaryotes, Argonaute proteins, guided by short RNA sequences, defend cells against transposons and viruses. In the eubacterium Thermus thermophilus, the DNA-guided Argonaute TtAgo defends against transformation by DNA plasmids. Here, we report that TtAgo also participates in DNA replication. In vivo, TtAgo binds 15- to 18-nt DNA guides derived from the chromosomal region where replication terminates and associates with proteins known to act in DNA replication. When gyrase, the sole T. thermophilus type II topoisomerase, is inhibited, TtAgo allows the bacterium to finish replicating its circular genome. In contrast, loss of gyrase and TtAgo activity slows growth and produces long sausage-like filaments in which the individual bacteria are linked by DNA. Finally, wild-type T. thermophilus outcompetes an otherwise isogenic strain lacking TtAgo. We propose that the primary role of TtAgo is to help T. thermophilus disentangle the catenated circular chromosomes generated by DNA replication.
Keywords: Argonaute; DNA replication; RNA silencing; Thermus thermophilus; TtAgo; decatenation; gyrase; pAGO; terminus of replication; topoisomerase.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests P.D.Z., S.M.J., and H.Z. have submitted a patent application regarding novel uses of TtAgo (PCT/US2016/025724).
Figures





Comment in
-
Prokaryotic Argonautes Function beyond Immunity by Unlinking Replicating Chromosomes.Cell. 2020 Sep 17;182(6):1381-1383. doi: 10.1016/j.cell.2020.08.037. Cell. 2020. PMID: 32946780
References
-
- Aubry A, Fisher LM, Jarlier V, and Cambau E (2006). First functional characterization of a singly expressed bacterial type II topoisomerase: the enzyme from Mycobacterium tuberculosis. Biochem Biophys Res Commun 348, 158–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
