

Antibiotic Resistance in Vibrio cholerae: Mechanistic Insights from IncC Plasmid-Mediated Dissemination of a Novel Family of Genomic Islands Inserted at trmE
- PMID: 32848007
- PMCID: PMC7449626
- DOI: 10.1128/mSphere.00748-20
Antibiotic Resistance in Vibrio cholerae: Mechanistic Insights from IncC Plasmid-Mediated Dissemination of a Novel Family of Genomic Islands Inserted at trmE
Abstract
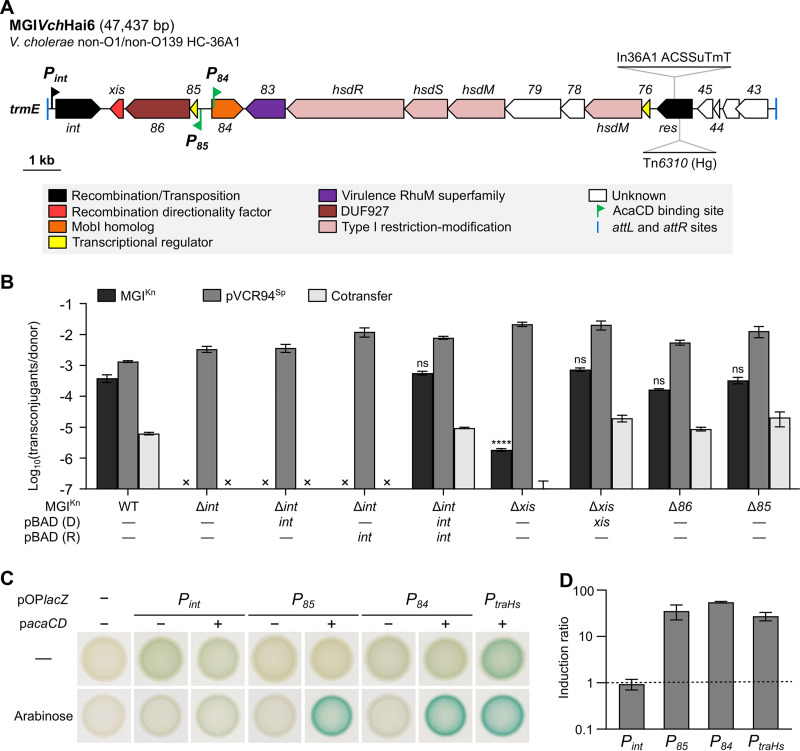
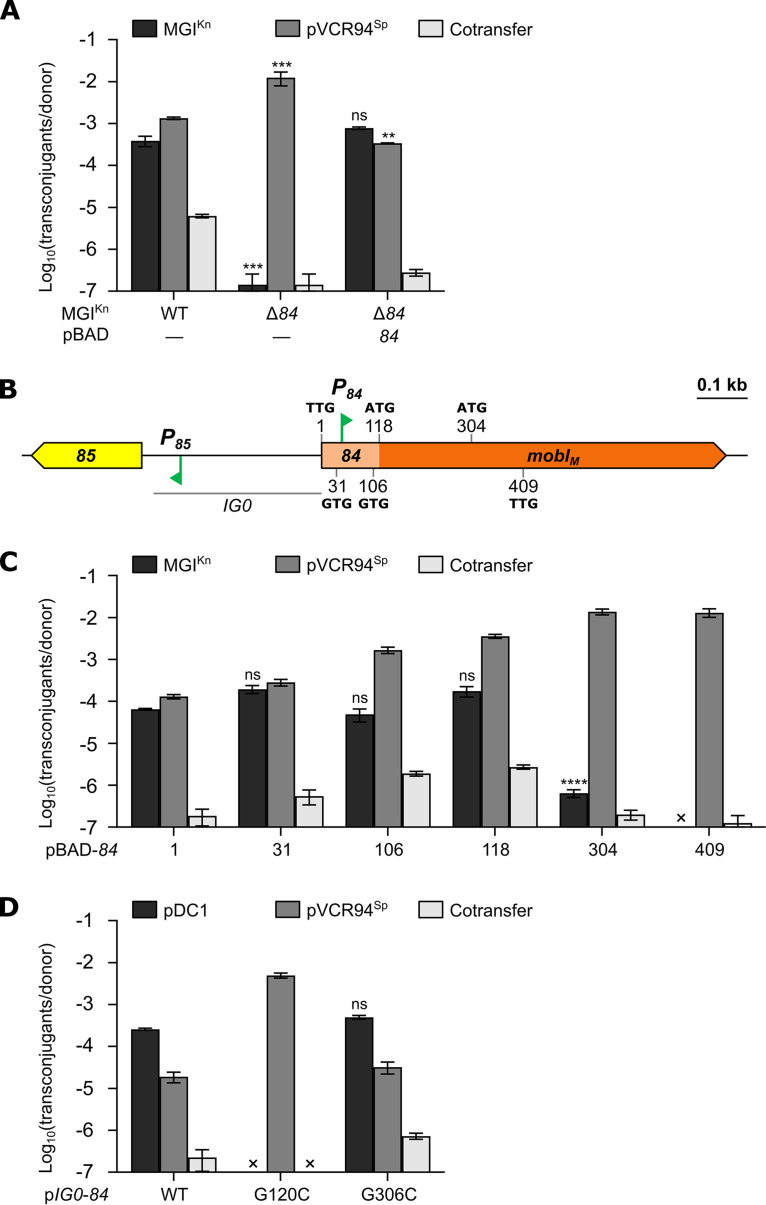
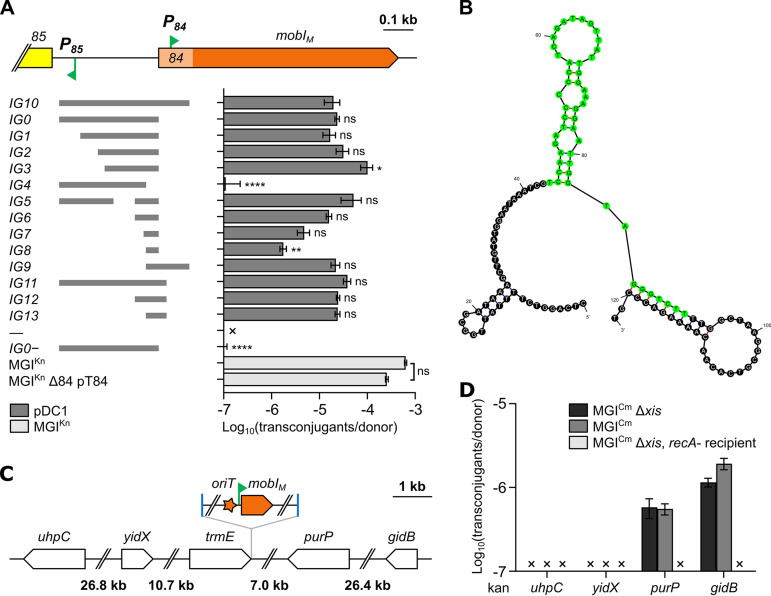
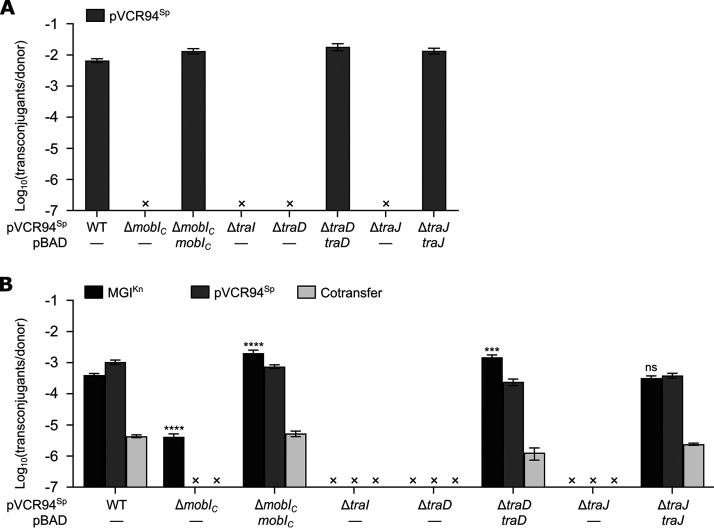
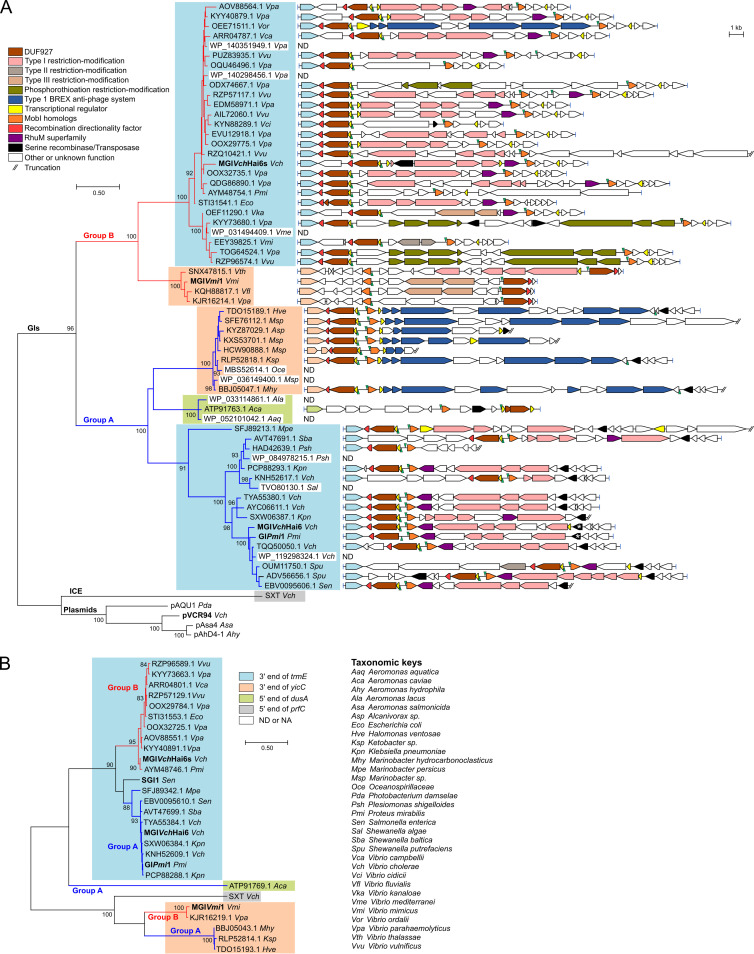
Cholera remains a formidable disease, and reports of multidrug-resistant strains of the causative agent Vibrio cholerae have become common during the last 3 decades. The pervasiveness of resistance determinants has largely been ascribed to mobile genetic elements, including SXT/R391 integrative conjugative elements, IncC plasmids, and genomic islands (GIs). Conjugative transfer of IncC plasmids is activated by the master activator AcaCD whose regulatory network extends to chromosomally integrated GIs. MGIVchHai6 is a multidrug resistance GI integrated at the 3' end of trmE (mnmE or thdF) in chromosome 1 of non-O1/non-O139 V. cholerae clinical isolates from the 2010 Haitian cholera outbreak. In the presence of an IncC plasmid expressing AcaCD, MGIVchHai6 excises from the chromosome and transfers at high frequency. Herein, the mechanism of mobilization of MGIVchHai6 GIs by IncC plasmids was dissected. Our results show that AcaCD drives expression of GI-borne genes, including xis and mobIM , involved in excision and mobilization. A 49-bp fragment upstream of mobIM was found to serve as the minimal origin of transfer (oriT) of MGIVchHai6. The direction of transfer initiated at oriT was determined using IncC plasmid-driven mobilization of chromosomal markers via MGIVchHai6. In addition, IncC plasmid-encoded factors, including the relaxase TraI, were found to be required for GI transfer. Finally, in silico exploration of Gammaproteobacteria genomes identified 47 novel related and potentially AcaCD-responsive GIs in 13 different genera. Despite sharing conserved features, these GIs integrate at trmE, yicC, or dusA and carry a diverse cargo of genes involved in phage resistance.IMPORTANCE The increasing association of the etiological agent of cholera, Vibrio cholerae serogroup O1 and O139, with multiple antibiotic resistance threatens to deprive health practitioners of this effective tool. Drug resistance in cholera results mainly from acquisition of mobile genetic elements. Genomic islands conferring multidrug resistance and mobilizable by IncC conjugative plasmids were reported to circulate in non-O1/non-O139 V. cholerae clinical strains isolated from the 2010 Haitian cholera outbreak. As these genomic islands can be transmitted to pandemic V. cholerae serogroups, their mechanism of transmission needed to be investigated. Our research revealed plasmid- and genomic island-encoded factors required for the resistance island excision, mobilization, and integration, as well as regulation of these functions. The discovery of related genomic islands carrying diverse phage resistance genes but lacking antibiotic resistance-conferring genes in a wide range of marine dwelling bacteria suggests that these elements are ancient and recently acquired drug resistance genes.
Keywords: IncC plasmids; T4CP; T4SS; Vibrio cholerae; antibiotic resistance; conjugation; genomic islands; horizontal gene transfer; mobilization; oriT; phage resistance; relaxase.
Copyright © 2020 Rivard et al.
Figures
References
-
- Verma J, Bag S, Saha B, Kumar P, Ghosh TS, Dayal M, Senapati T, Mehra S, Dey P, Desigamani A, Kumar D, Rana P, Kumar B, Maiti TK, Sharma NC, Bhadra RK, Mutreja A, Nair GB, Ramamurthy T, Das B. 2019. Genomic plasticity associated with antimicrobial resistance in Vibrio cholerae. Proc Natl Acad Sci U S A 116:6226–6231. doi:10.1073/pnas.1900141116. - DOI - PMC - PubMed
-
- Weill F-X, Domman D, Njamkepo E, Tarr C, Rauzier J, Fawal N, Keddy KH, Salje H, Moore S, Mukhopadhyay AK, Bercion R, Luquero FJ, Ngandjio A, Dosso M, Monakhova E, Garin B, Bouchier C, Pazzani C, Mutreja A, Grunow R, Sidikou F, Bonte L, Breurec S, Damian M, Njanpop-Lafourcade B-M, Sapriel G, Page A-L, Hamze M, Henkens M, Chowdhury G, Mengel M, Koeck J-L, Fournier J-M, Dougan G, Grimont PAD, Parkhill J, Holt KE, Piarroux R, Ramamurthy T, Quilici M-L, Thomson NR. 2017. Genomic history of the seventh pandemic of cholera in Africa. Science 358:785–789. doi:10.1126/science.aad5901. - DOI - PubMed
-
- Spagnoletti M, Ceccarelli D, Rieux A, Fondi M, Taviani E, Fani R, Colombo MM, Colwell RR, Balloux F. 2014. Acquisition and evolution of SXT-R391 integrative conjugative elements in the seventh-pandemic Vibrio cholerae lineage. mBio 5:e01356-14. [CrossRef] doi:10.1128/mBio.01356-14. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
