

Post-translational modification as a response to cellular stress induced by hemoglobin oxidation in sickle cell disease
- PMID: 32848178
- PMCID: PMC7450072
- DOI: 10.1038/s41598-020-71096-6
Post-translational modification as a response to cellular stress induced by hemoglobin oxidation in sickle cell disease
Abstract
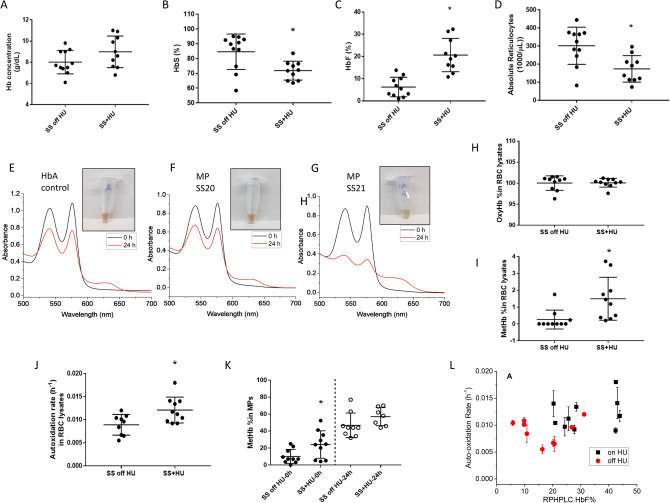
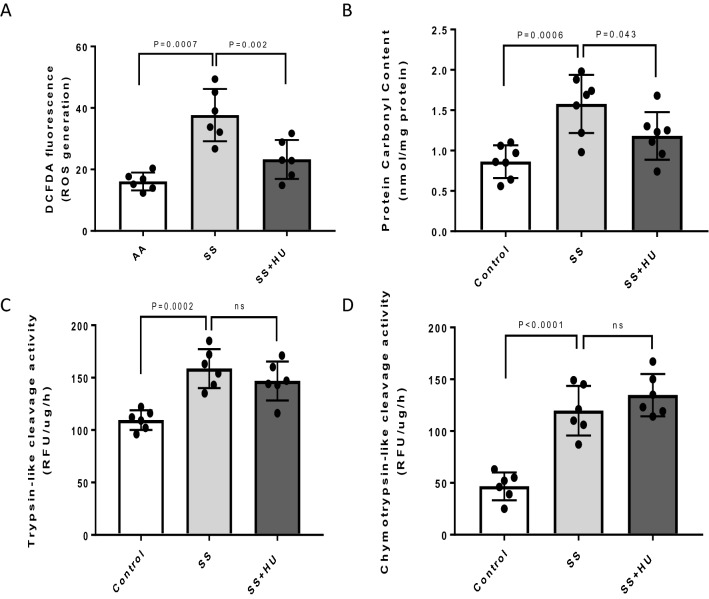
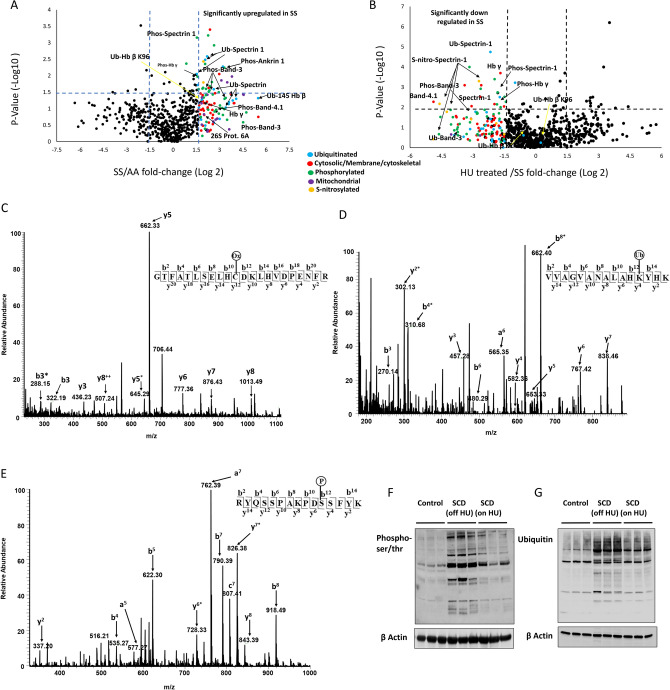
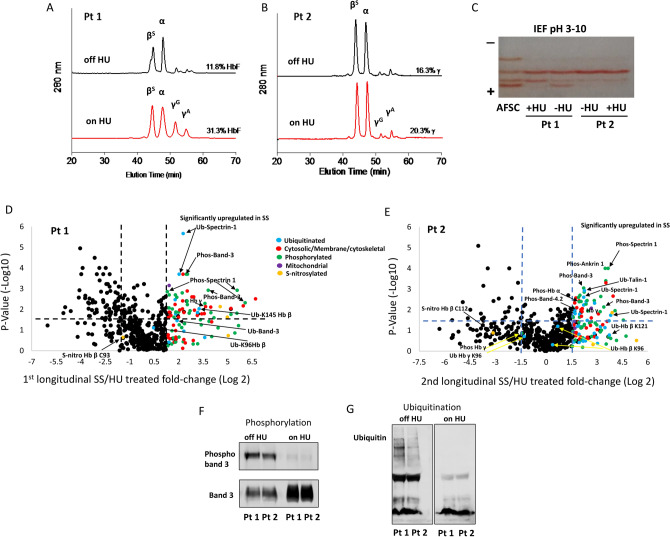
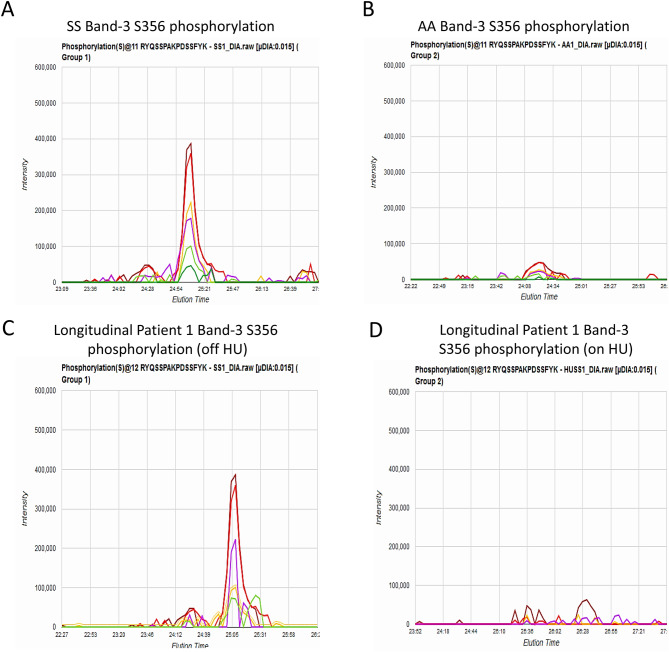
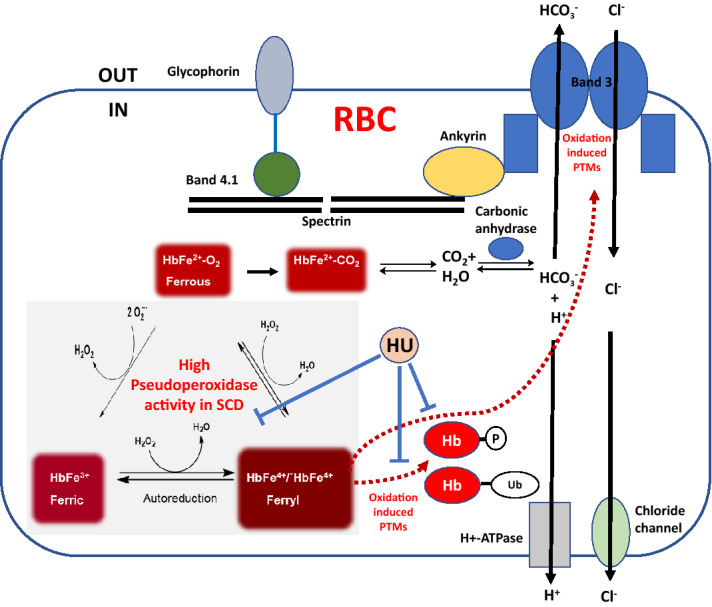
Intracellular oxidative stress and oxidative modification of sickle hemoglobin (HbS) play a role in sickle cell disease (SCD) pathogenesis. Recently, we reported that Hb-dependent oxidative stress induced post-translational modifications (PTMs) of Hb and red blood cell (RBC) membrane proteins of transgenic SCD mice. To identify the mechanistic basis of these protein modifications, we followed in vitro oxidative changes occurring in intracellular Hb obtained from RBCs and RBC-derived microparticles (MPs) from the blood of 23 SCD patients (HbSS) of which 11 were on, and 12, off hydroxyurea (HU) treatment, and 5 ethnic matched controls. We used mass spectrometry-based proteomics to characterize these oxidative PTMs on a cross-sectional group of these patients (n = 4) and a separate subgroup of patients (n = 2) studied prior to initiation and during HU treatment. Collectively, these data indicated that band-3 and its interaction network involved in MPs formation exhibited more protein phosphorylation and ubiquitination in SCD patients than in controls. HU treatment reversed these oxidative PTMs back to level observed in controls. These PTMs were also confirmed using orthogonal immunoprecipitation experiments. Moreover, we observed specific markers reflective of oxidative stress, including irreversible oxidation of βCys93 and ubiquitination of Hb βLys145 (and βLys96). Overall, these studies strongly suggest that extensive erythrocyte membrane protein phosphorylation and ubiquitination are involved in SCD pathogenesis and provide further insight into the multifaceted effects of HU treatment.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
