

Ketotherapeutics for neurodegenerative diseases
- PMID: 32854853
- PMCID: PMC7418707
- DOI: 10.1016/bs.irn.2020.02.003
Ketotherapeutics for neurodegenerative diseases
Abstract
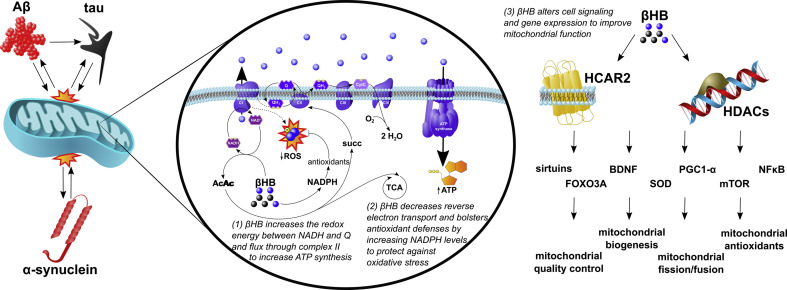
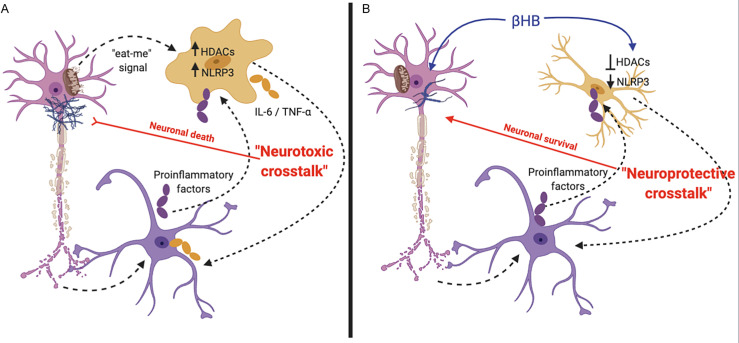
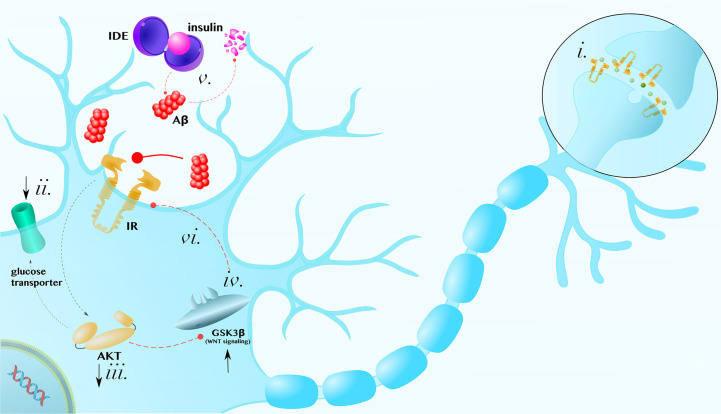
Alzheimer's disease (AD) and Parkinson's disease (PD) are, respectively, the most prevalent and fastest growing neurodegenerative diseases worldwide. The former is primarily characterized by memory loss and the latter by the motor symptoms of tremor and bradykinesia. Both AD and PD are progressive diseases that share several key underlying mitochondrial, inflammatory, and other metabolic pathologies. This review will detail how these pathologies intersect with ketone body metabolism and signaling, and how ketone bodies, particularly d-β-hydroxybutyrate (βHB), may serve as a potential adjunctive nutritional therapy for two of the world's most devastating conditions.
Keywords: Alzheimer's disease; Inflammation; Insulin; Ketone bodies; Ketone ester; Microglia; Mitochondria; Oxidative stress; Parkinson's disease; d-β-hydroxybutyrate.
© 2020 Elsevier Inc. All rights reserved.
Figures
References
-
- Andersen J.V., Christensen S.K., Aldana B.I., Nissen J.D., Tanila K., Waagepetersen H.S. Alterations in cerebral cortical glucose and glutamine metabolism precedes amyloid plaques in the APPswe/PSEN1dE9 mouse model of Alzheimer's disease. Neurochemical Research. 2017;42:1589–1598. doi: 10.1007/s11064-016-2070-2. - DOI - PubMed
Further reading
-
- Chriett S., Dąbek A., Wojtala M., Vidal H., Balcerczyk A., Pirola L. Prominent action of butyrate over β-hydroxybutyrate as histone deacetylase inhibitor, transcriptional modulator and anti-inflammatory molecule. Scientific Reports. 2019;9(1):1–14. doi: 10.1038/s41598-018-36941-9. - DOI - PMC - PubMed
-
- Deora V., Albornoz E.A., Zhu K., Woodruff T.M., Gordon R. The ketone body β-hydroxybutyrate does not inhibit synuclein mediated inflammasome activation in microglia. Journal of Neuroimmune Pharmacology: The Official Journal of the Society on NeuroImmune Pharmacology. 2017;12(4):568–574. doi: 10.1007/s11481-017-9754-5. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
