

Single cell transcriptomics identifies a signaling network coordinating endoderm and mesoderm diversification during foregut organogenesis
- PMID: 32855417
- PMCID: PMC7453027
- DOI: 10.1038/s41467-020-17968-x
Single cell transcriptomics identifies a signaling network coordinating endoderm and mesoderm diversification during foregut organogenesis
Abstract
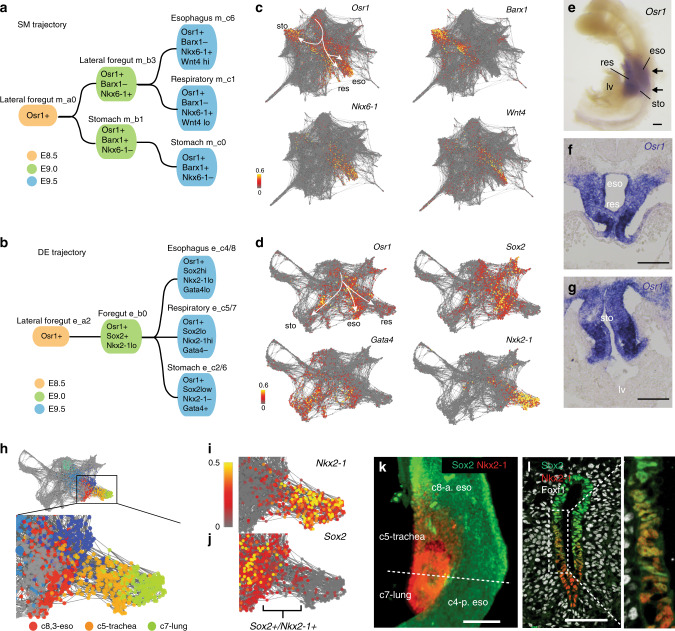
Visceral organs, such as the lungs, stomach and liver, are derived from the fetal foregut through a series of inductive interactions between the definitive endoderm (DE) and the surrounding splanchnic mesoderm (SM). While DE patterning is fairly well studied, the paracrine signaling controlling SM regionalization and how this is coordinated with epithelial identity is obscure. Here, we use single cell transcriptomics to generate a high-resolution cell state map of the embryonic mouse foregut. This identifies a diversity of SM cell types that develop in close register with the organ-specific epithelium. We infer a spatiotemporal signaling network of endoderm-mesoderm interactions that orchestrate foregut organogenesis. We validate key predictions with mouse genetics, showing the importance of endoderm-derived signals in mesoderm patterning. Finally, leveraging these signaling interactions, we generate different SM subtypes from human pluripotent stem cells (hPSCs), which previously have been elusive. The single cell data can be explored at: https://research.cchmc.org/ZornLab-singlecell .
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Le Douarin N, Houssaint E. Role of the mesoderm in the induction of the synthesis of glycogen during differentiation of the hepatic endoderm. C. R. Acad. Hebd. Seances Acad. Sci. D. 1967;264:1872–1874. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
