

The T-type calcium channel isoform Cav3.1 is a target for the hypnotic effect of the anaesthetic neurosteroid (3β,5β,17β)-3-hydroxyandrostane-17-carbonitrile
- PMID: 32859366
- PMCID: PMC7844375
- DOI: 10.1016/j.bja.2020.07.022
The T-type calcium channel isoform Cav3.1 is a target for the hypnotic effect of the anaesthetic neurosteroid (3β,5β,17β)-3-hydroxyandrostane-17-carbonitrile
Abstract
Background: The mechanisms underlying the role of T-type calcium channels (T-channels) in thalamocortical excitability and oscillations in vivo during neurosteroid-induced hypnosis are largely unknown.
Methods: We used patch-clamp electrophysiological recordings from acute brain slices ex vivo, recordings of local field potentials (LFPs) from the central medial thalamic nucleus in vivo, and wild-type (WT) and Cav3.1 knock-out mice to investigate the molecular mechanisms of hypnosis induced by the neurosteroid analogue (3β,5β,17β)-3-hydroxyandrostane-17-carbonitrile (3β-OH).
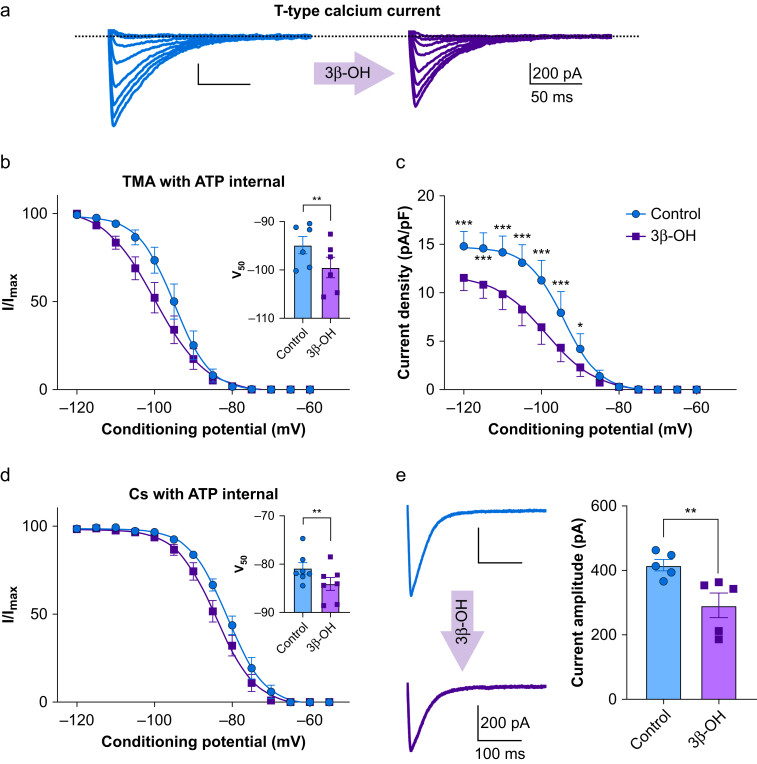
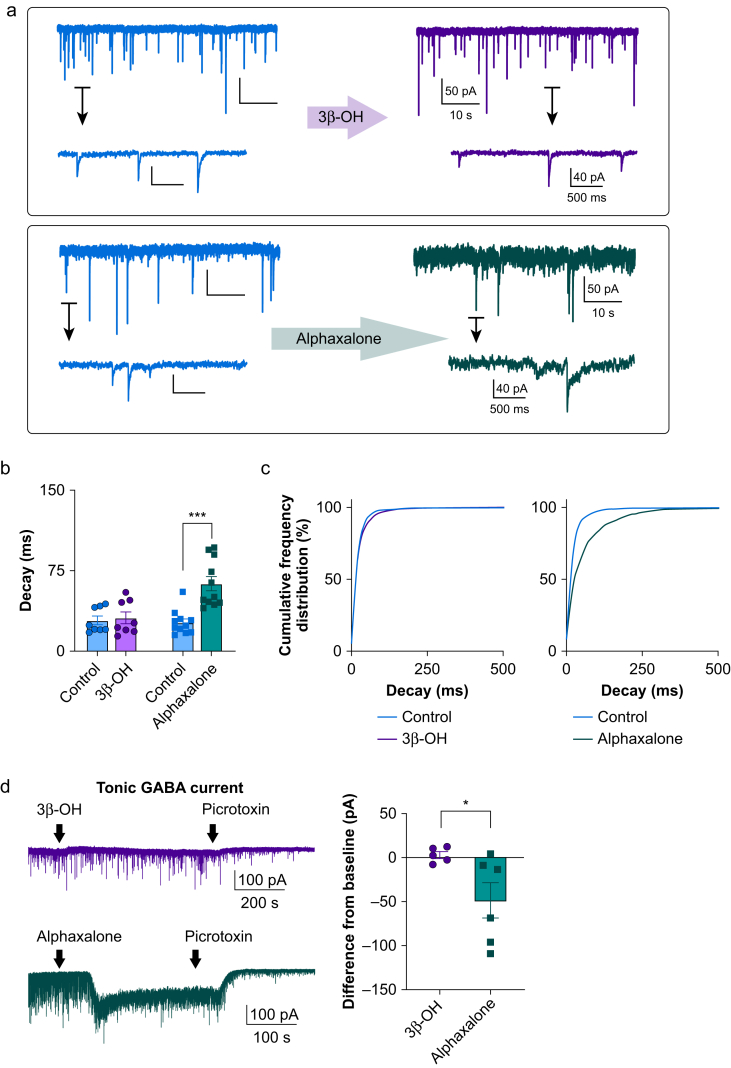
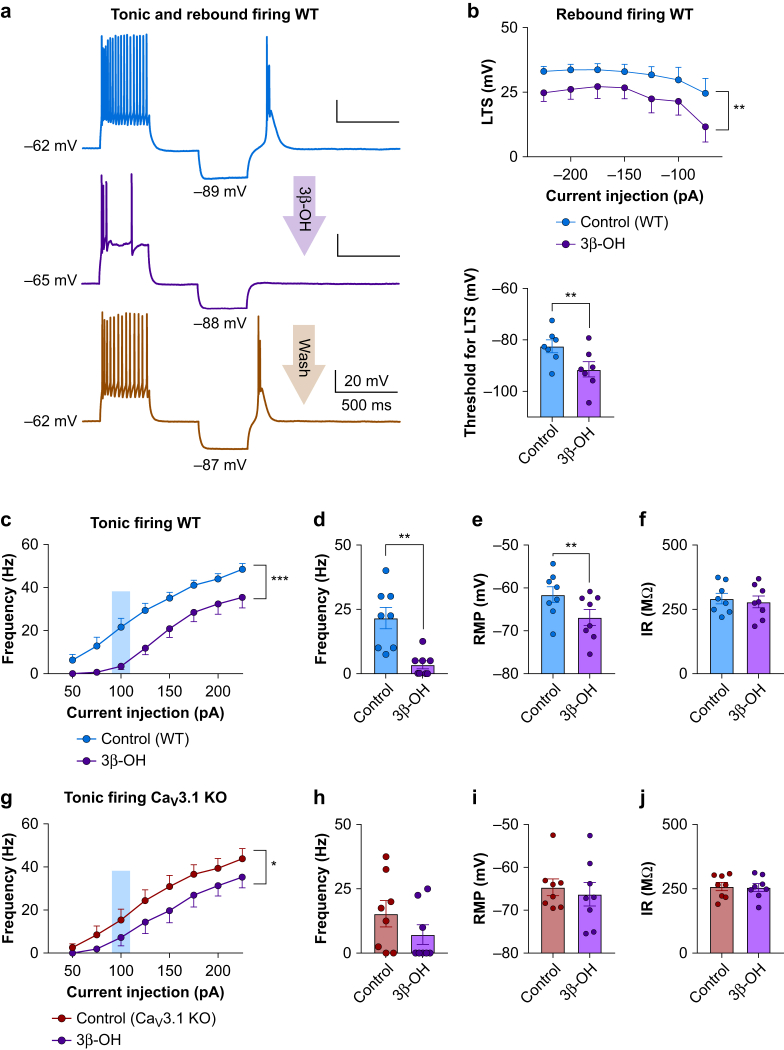
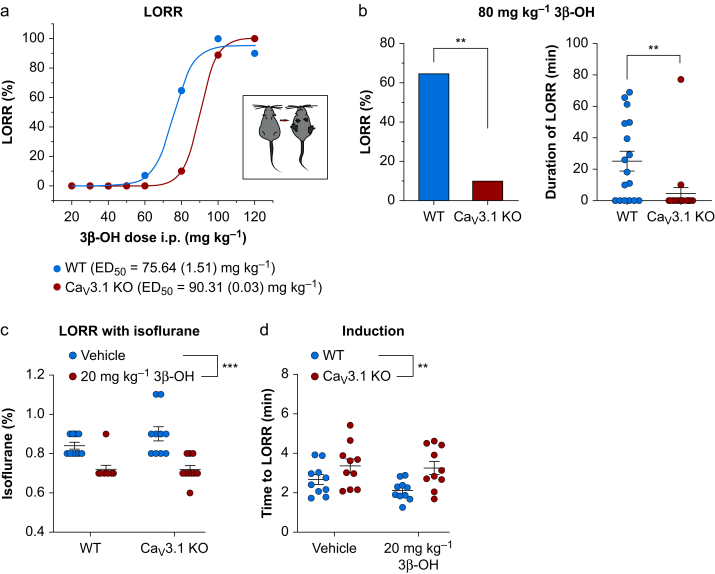
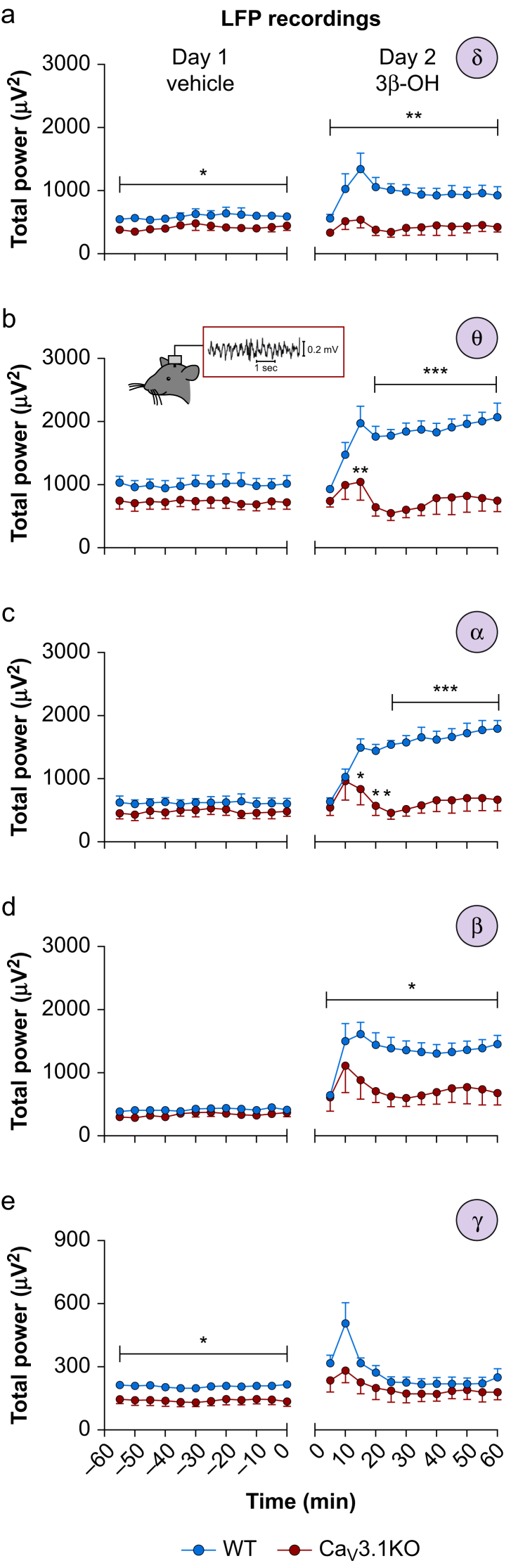
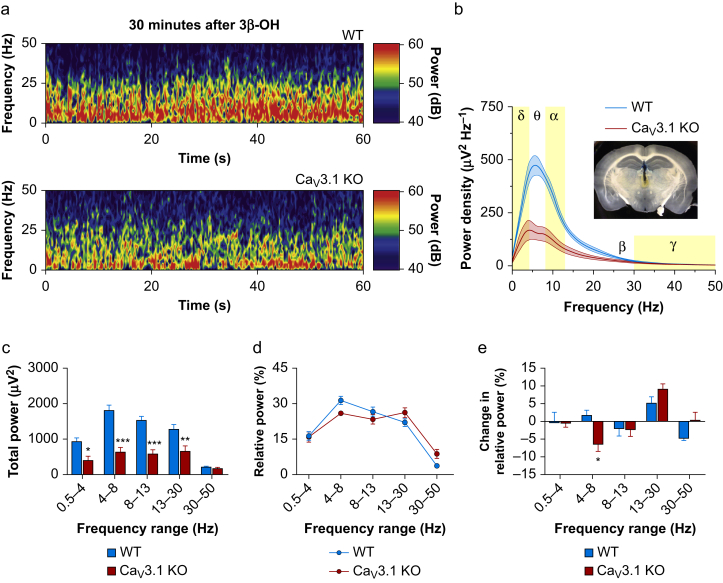
Results: Patch-clamp recordings showed that 3β-OH inhibited isolated T-currents but had no effect on phasic or tonic γ-aminobutyric acid A currents. Also in acute brain slices, 3β-OH inhibited the spike firing mode more profoundly in WT than in Cav3.1 knockout mice. Furthermore, 3β-OH significantly hyperpolarised neurones, reduced the amplitudes of low threshold spikes, and diminished rebound burst firing only in WT mice. We found that 80 mg kg-1 i.p. injections of 3β-OH induced hypnosis in >60% of WT mice but failed to induce hypnosis in the majority of mutant mice. A subhypnotic dose of 3β-OH (20 mg kg-1 i.p.) accelerated induction of hypnosis by isoflurane only in WT mice, but had similar effects on the maintenance of isoflurane-induced hypnosis in both WT and Cav3.1 knockout mice. In vivo recordings of LFPs showed that a hypnotic dose of 3β-OH increased δ, θ, α, and β oscillations in WT mice in comparison with Cav3.1 knock-out mice.
Conclusions: The Cav3.1 T-channel isoform is critical for diminished thalamocortical excitability and oscillations that underlie neurosteroid-induced hypnosis.
Keywords: calcium channel; electrophysiology; hypnosis; mechanisms of anaesthesia; neurosteroid; thalamus.
Copyright © 2020 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Anaesthetic-induced developmental neurotoxicity on (neuro)steroids.Br J Anaesth. 2021 Jan;126(1):34-37. doi: 10.1016/j.bja.2020.08.008. Epub 2020 Sep 3. Br J Anaesth. 2021. PMID: 32891411 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
