

Low-dimensional dynamics for working memory and time encoding
- PMID: 32859756
- PMCID: PMC7502752
- DOI: 10.1073/pnas.1915984117
Low-dimensional dynamics for working memory and time encoding
Erratum in
-
Correction for Cueva et al., Low-dimensional dynamics for working memory and time encoding.Proc Natl Acad Sci U S A. 2023 Mar 14;120(11):e2302618120. doi: 10.1073/pnas.2302618120. Epub 2023 Mar 10. Proc Natl Acad Sci U S A. 2023. PMID: 36897990 Free PMC article. No abstract available.
Abstract
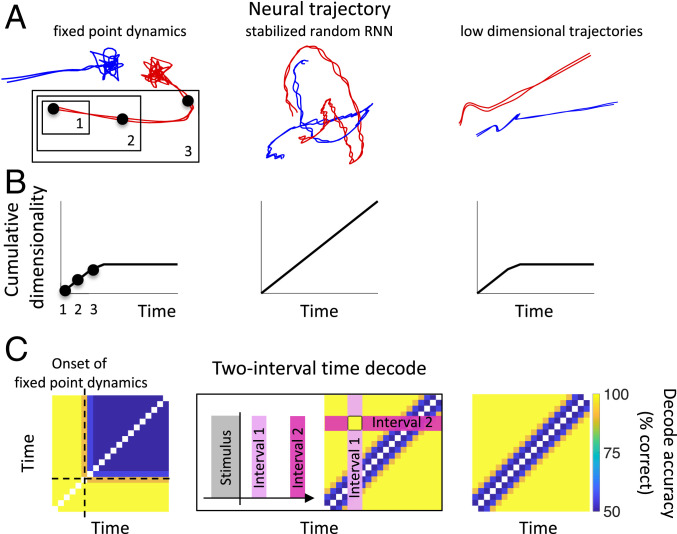
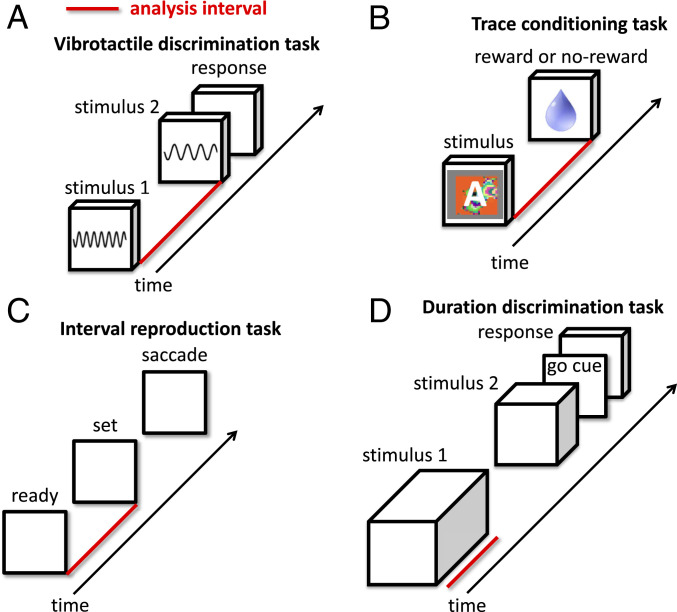
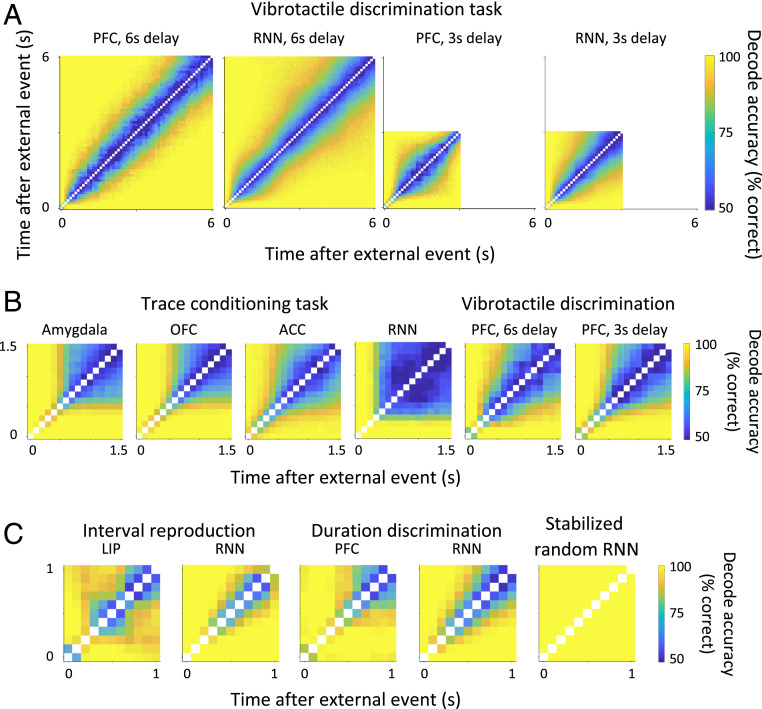
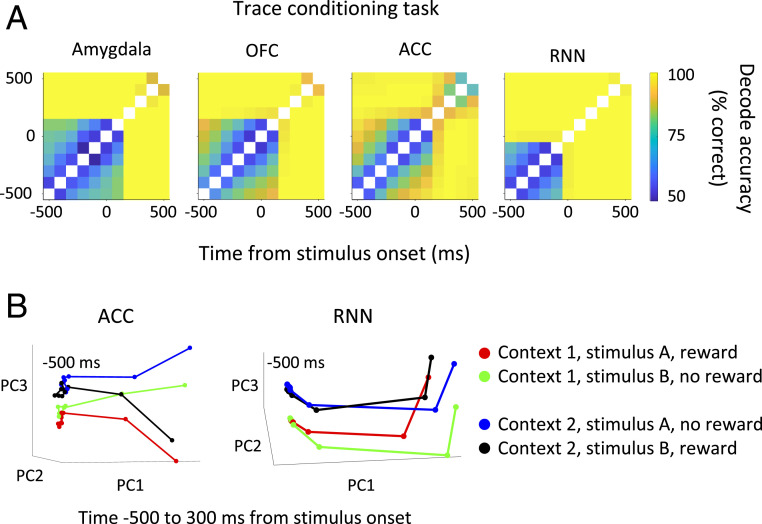
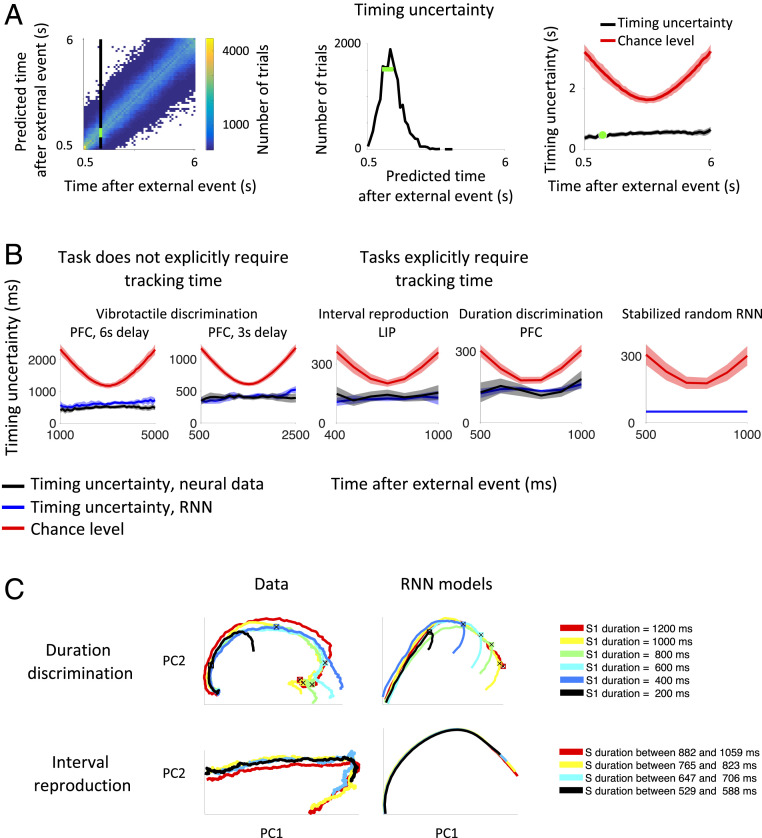
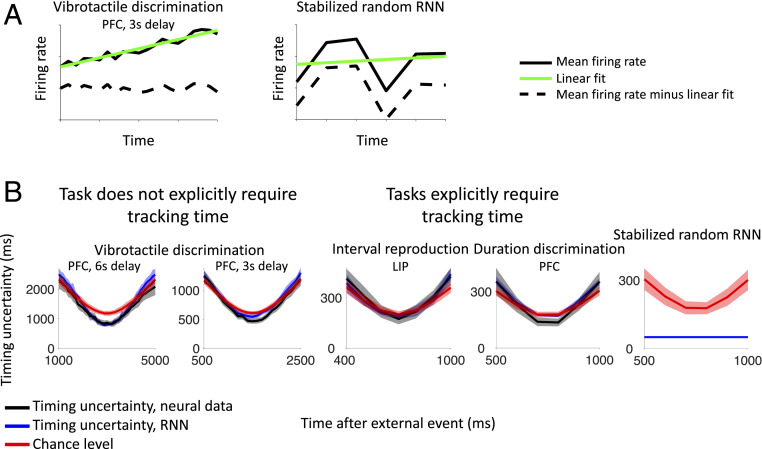
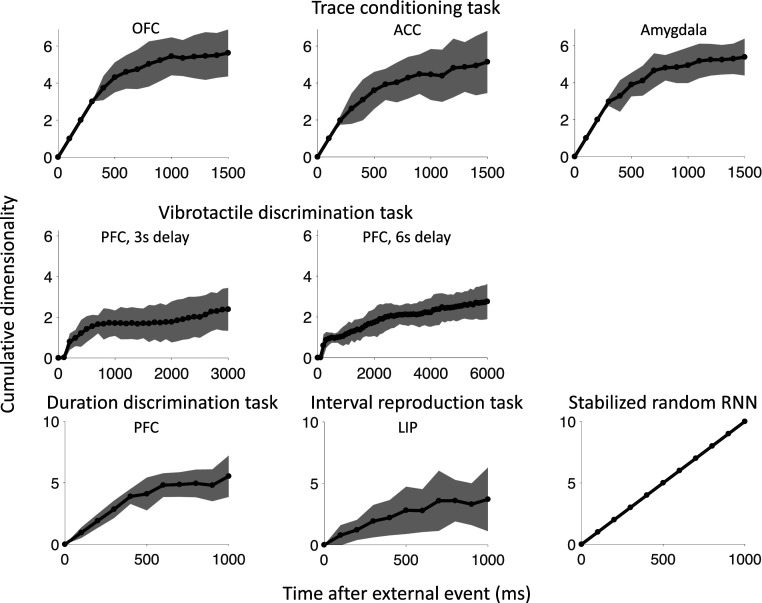
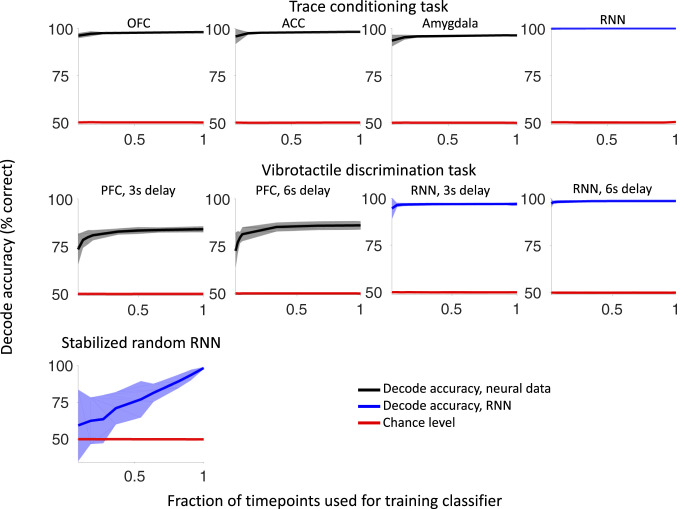
Our decisions often depend on multiple sensory experiences separated by time delays. The brain can remember these experiences and, simultaneously, estimate the timing between events. To understand the mechanisms underlying working memory and time encoding, we analyze neural activity recorded during delays in four experiments on nonhuman primates. To disambiguate potential mechanisms, we propose two analyses, namely, decoding the passage of time from neural data and computing the cumulative dimensionality of the neural trajectory over time. Time can be decoded with high precision in tasks where timing information is relevant and with lower precision when irrelevant for performing the task. Neural trajectories are always observed to be low-dimensional. In addition, our results further constrain the mechanisms underlying time encoding as we find that the linear "ramping" component of each neuron's firing rate strongly contributes to the slow timescale variations that make decoding time possible. These constraints rule out working memory models that rely on constant, sustained activity and neural networks with high-dimensional trajectories, like reservoir networks. Instead, recurrent networks trained with backpropagation capture the time-encoding properties and the dimensionality observed in the data.
Keywords: neural dynamics; recurrent networks; reservoir computing; time decoding; working memory.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Baddeley A., Hitch G., “Working memory” in Psychology of Learning and Motivation Bower G. A., Ed. (Academic, New York, 1974), vol. 8, pp. 47–90.
-
- Miyake A., Shah P., Models of Working Memory (Cambridge University Press, 1999).
-
- Gibbon J., Malapani C., Dale C. L., Gallistel C. R., Toward a neurobiology of temporal cognition: Advances and challenges. Curr. Opin. Neurobiol. 7, 170–184 (1997). - PubMed
-
- Buonomano D. V., Karmarkar U. R., How do we tell time? Neuroscientist 8, 42–51 (2002). - PubMed
-
- Amit D. J., Modeling Brain Function: The World of Attractor Neural Networks (Cambridge University Press, 1992).
