

A pathogenic and clonally expanded B cell transcriptome in active multiple sclerosis
- PMID: 32859762
- PMCID: PMC7502747
- DOI: 10.1073/pnas.2008523117
A pathogenic and clonally expanded B cell transcriptome in active multiple sclerosis
Abstract
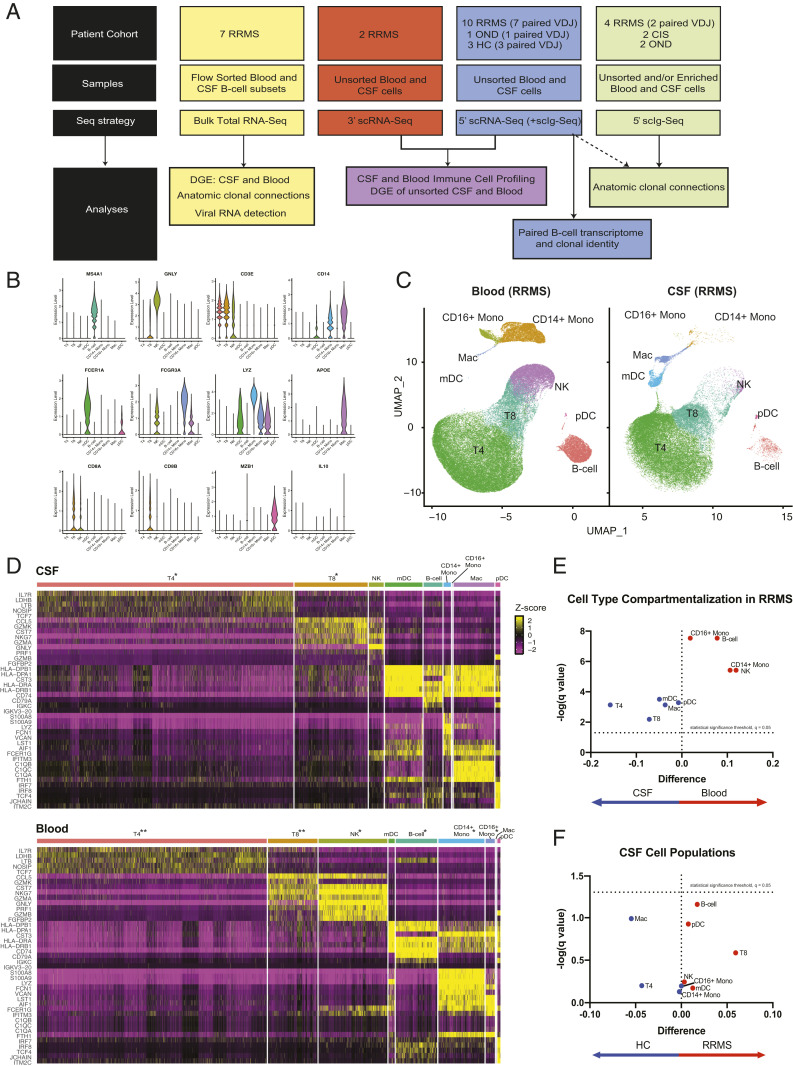
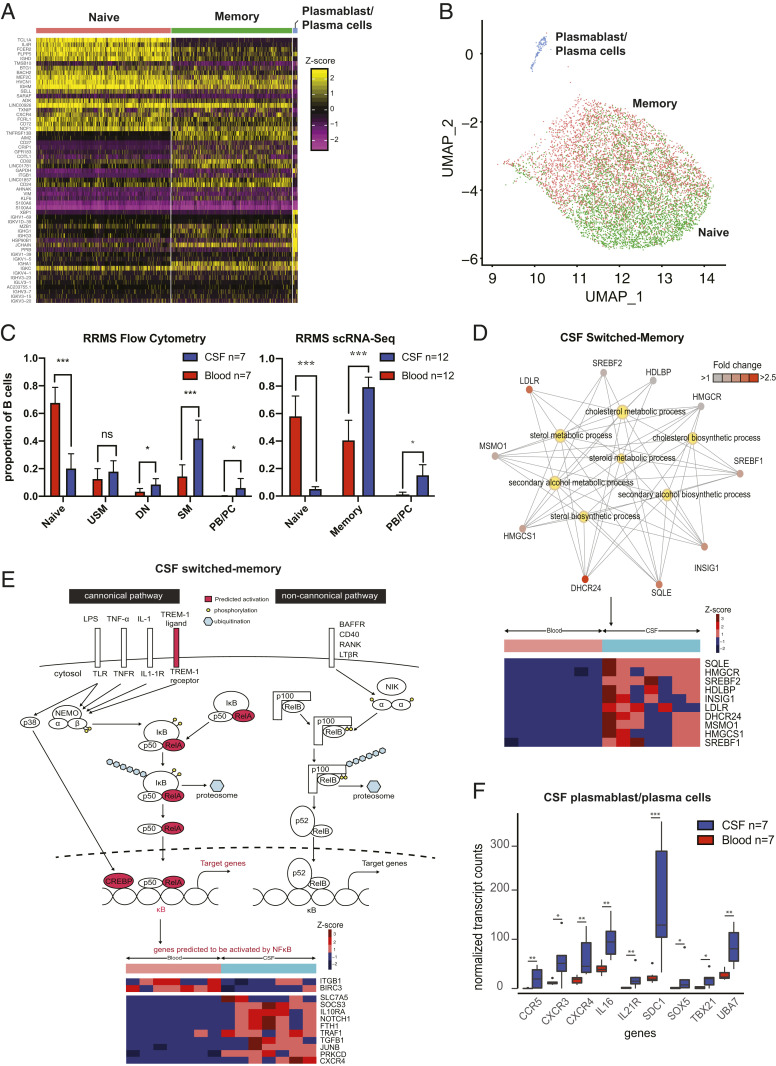
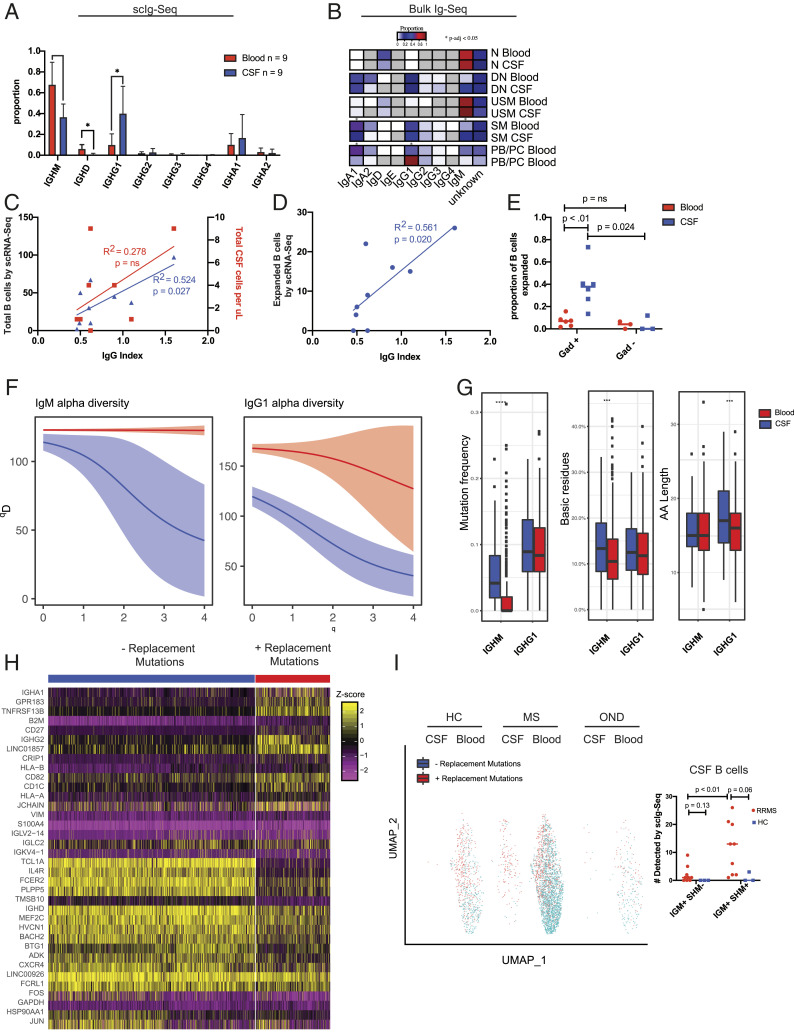
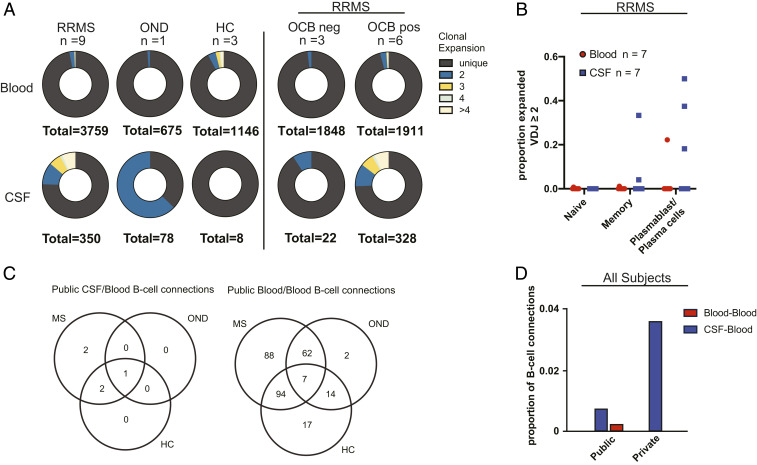
Central nervous system B cells have several potential roles in multiple sclerosis (MS): secretors of proinflammatory cytokines and chemokines, presenters of autoantigens to T cells, producers of pathogenic antibodies, and reservoirs for viruses that trigger demyelination. To interrogate these roles, single-cell RNA sequencing (scRNA-Seq) was performed on paired cerebrospinal fluid (CSF) and blood from subjects with relapsing-remitting MS (RRMS; n = 12), other neurologic diseases (ONDs; n = 1), and healthy controls (HCs; n = 3). Single-cell immunoglobulin sequencing (scIg-Seq) was performed on a subset of these subjects and additional RRMS (n = 4), clinically isolated syndrome (n = 2), and OND (n = 2) subjects. Further, paired CSF and blood B cell subsets (RRMS; n = 7) were isolated using fluorescence activated cell sorting for bulk RNA sequencing (RNA-Seq). Independent analyses across technologies demonstrated that nuclear factor kappa B (NF-κB) and cholesterol biosynthesis pathways were activated, and specific cytokine and chemokine receptors were up-regulated in CSF memory B cells. Further, SMAD/TGF-β1 signaling was down-regulated in CSF plasmablasts/plasma cells. Clonally expanded, somatically hypermutated IgM+ and IgG1+ CSF B cells were associated with inflammation, blood-brain barrier breakdown, and intrathecal Ig synthesis. While we identified memory B cells and plasmablast/plasma cells with highly similar Ig heavy-chain sequences across MS subjects, similarities were also identified with ONDs and HCs. No viral transcripts, including from Epstein-Barr virus, were detected. Our findings support the hypothesis that in MS, CSF B cells are driven to an inflammatory and clonally expanded memory and plasmablast/plasma cell phenotype.
Keywords: B cell; immune repertoire; multiple sclerosis; neuroimmunology.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: R.D.S. served on an advisory board for Sanofi Genzyme and is currently an employee of Asceneuron SA. A.J.G. reports personal fees from Inception Sciences and Mylan Pharmaceuticals and has reported serving on an end point adjudication committee for Biogen and Medimmune. He has served on trial steering committees for Novartis and serves on the Scientific Advisory Board for Bionure. R.M.B. has received personal compensation for medical legal consulting and for consulting or serving on the advisory boards of F. Hoffmann-La Roche Ltd., Sanofi-Genzyme, and Novartis. J.M.G. has received research support to University of California, San Francisco from Genentech, personal compensation for consulting for Biogen and Alexion, and personal compensation for medical legal consulting; he has also received honoraria from Dynamed Plus for editorial work. B.A.C.C. receives personal compensation for consulting from Abbvie, Biogen, EMD Serono, GeNeuro, Novartis, and Sanofi Genzyme. S.S.Z. is Deputy Editor of Neurology, Neuroimmunology and Neuroinflammation and is an Associate Editor for Frontiers in Immunology and Frontiers in Neurology. He serves on the Advisory Committee for the American Congress on Treatment and Research in Multiple Sclerosis and on the grant review committee for the National Multiple Sclerosis Society (NMSS). Previously, he has served on the Editorial Board of the Journal of Clinical Investigation, The Journal of Immunology, and The Journal of Neurological Sciences and has been a charter member of the grant review committee for the NIH Clinical Neuroimmunology and Brain Tumors. He has served as a consultant and received honoraria from Biogen-Idec, EMD-Serono, Genzyme, Novartis, Roche/Genentech, and Teva Pharmaceuticals, Inc. and has served on Data Safety Monitoring Boards for Lilly, BioMS, Teva, and Opexa Therapeutics. Currently, S.S.Z. receives research grant support from the NIH, NMSS, Weill Institute, Race to Erase MS, and the Maisin Foundation. S.L.H. currently serves on the Scientific Advisory Board of Alector, Annexon, Bionure, and Molecular Stethoscope and on the Board of Trustees of Neurona. S.L.H. also has received travel reimbursement and writing assistance from F. Hoffmann-La Roche Ltd. and Novartis for CD20-related meetings and presentations. M.R.W. received research funding from Roche/Genentech.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
