

Structural and Biophysical Mechanisms of Class C G Protein-Coupled Receptor Function
- PMID: 32861513
- PMCID: PMC7642020
- DOI: 10.1016/j.tibs.2020.07.008
Structural and Biophysical Mechanisms of Class C G Protein-Coupled Receptor Function
Abstract
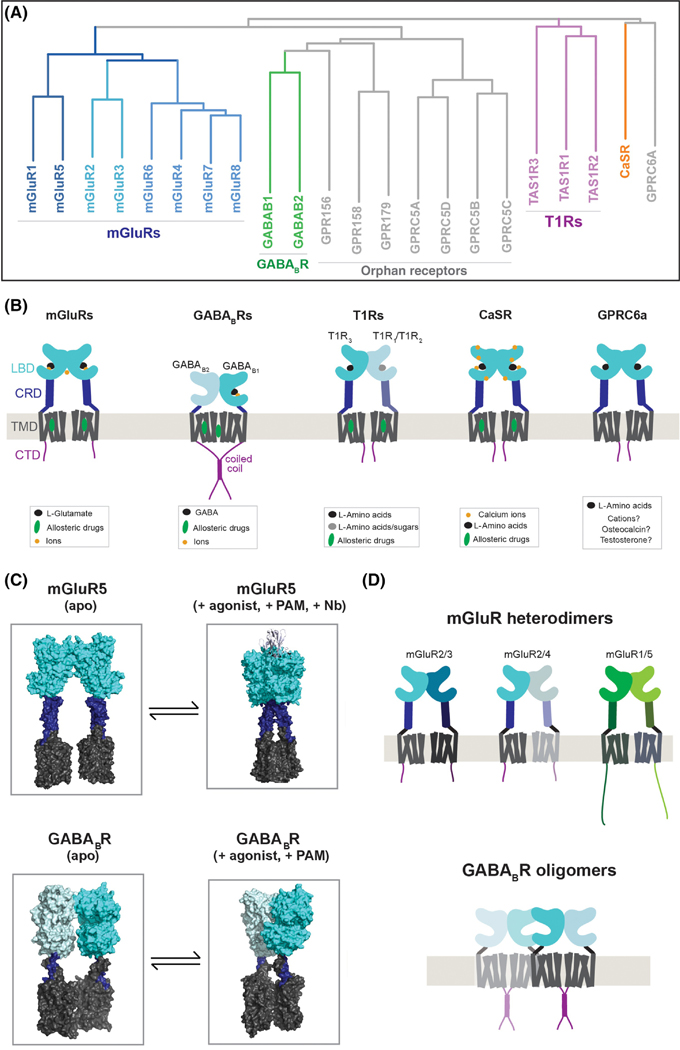
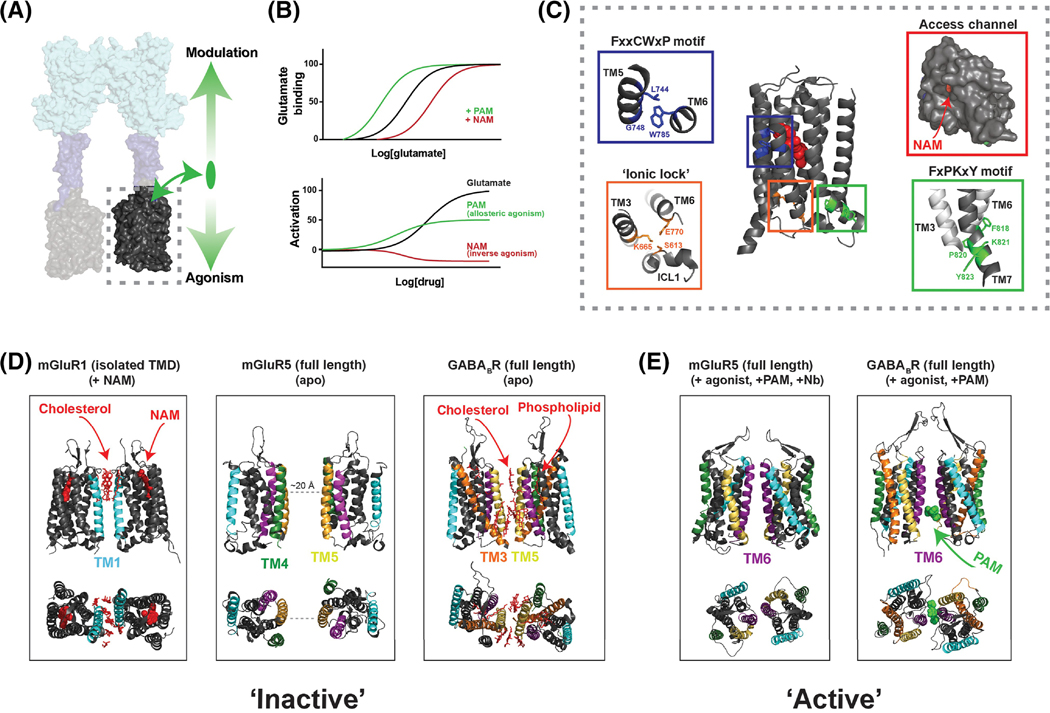
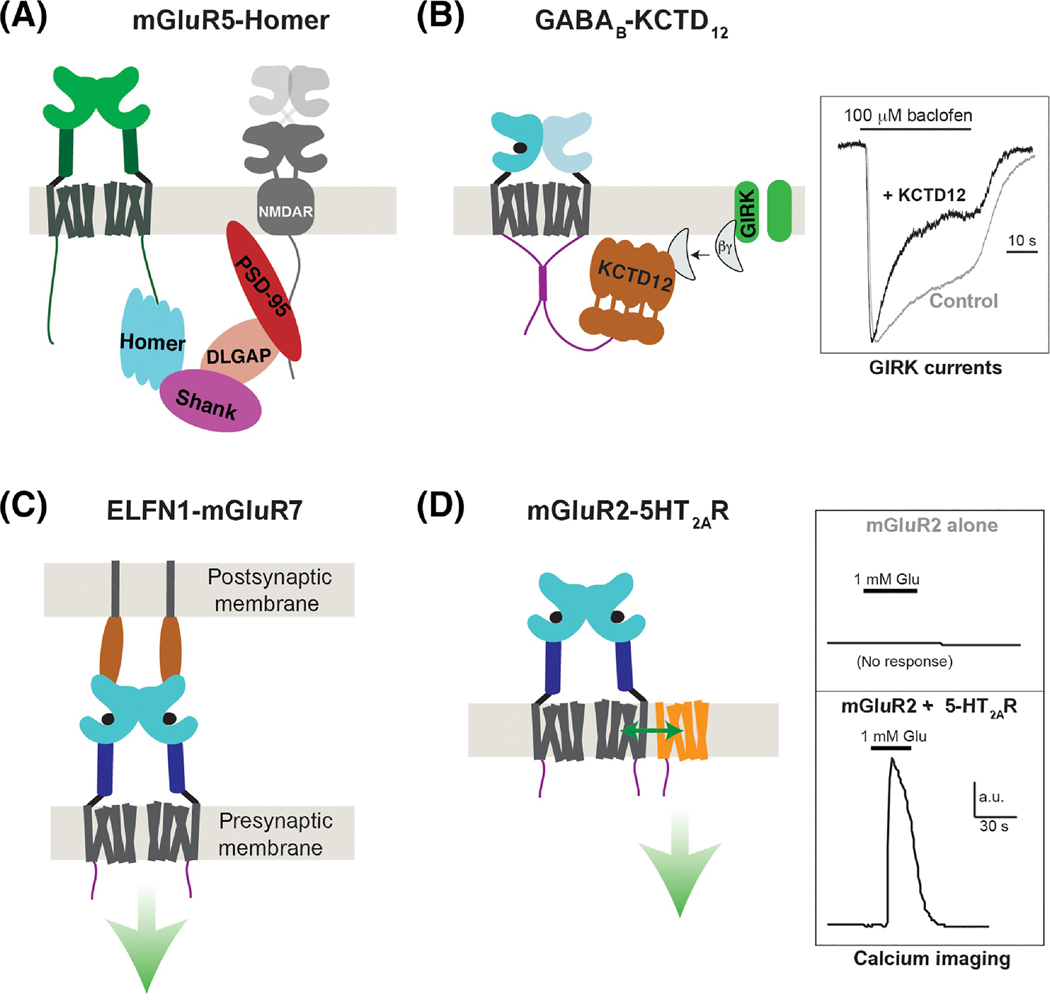
Groundbreaking structural and spectroscopic studies of class A G protein-coupled receptors (GPCRs), such as rhodopsin and the β2 adrenergic receptor, have provided a picture of how structural rearrangements between transmembrane helices control ligand binding, receptor activation, and effector coupling. However, the activation mechanism of other GPCR classes remains more elusive, in large part due to complexity in their domain assembly and quaternary structure. In this review, we focus on the class C GPCRs, which include metabotropic glutamate receptors (mGluRs) and gamma-aminobutyric acid B (GABAB) receptors (GABABRs) most prominently. We discuss the unique biophysical questions raised by the presence of large extracellular ligand-binding domains (LBDs) and constitutive homo/heterodimerization. Furthermore, we discuss how recent studies have begun to unravel how these fundamental class C GPCR features impact the processes of ligand binding, receptor activation, signal transduction, regulation by accessory proteins, and crosstalk with other GPCRs.
Keywords: G protein-coupled receptors; GABA(B) receptor; calcium-sensing receptor; metabotropic glutamate receptor.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures

References
-
- de Graaf C. et al. (2017) Extending the Structural View of Class B GPCRs. Trends Biochem Sci 42 (12), 946–960. - PubMed
-
- Pin JP and Bettler B. (2016) Organization and functions of mGlu and GABAB receptor complexes. Nature 540 (7631), 60–68. - PubMed
-
- Zhang X. et al. (2018) Structural and Druggability Landscape of Frizzled G Protein-Coupled Receptors. Trends Biochem Sci 43 (12), 1033–1046. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
