

Structure, function, and inhibitor targeting of HIV-1 Nef-effector kinase complexes
- PMID: 32862141
- PMCID: PMC7606690
- DOI: 10.1074/jbc.REV120.012317
Structure, function, and inhibitor targeting of HIV-1 Nef-effector kinase complexes
Abstract
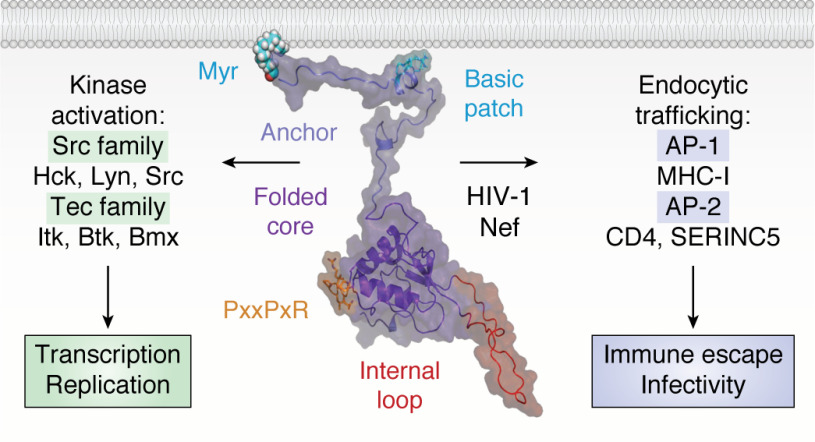
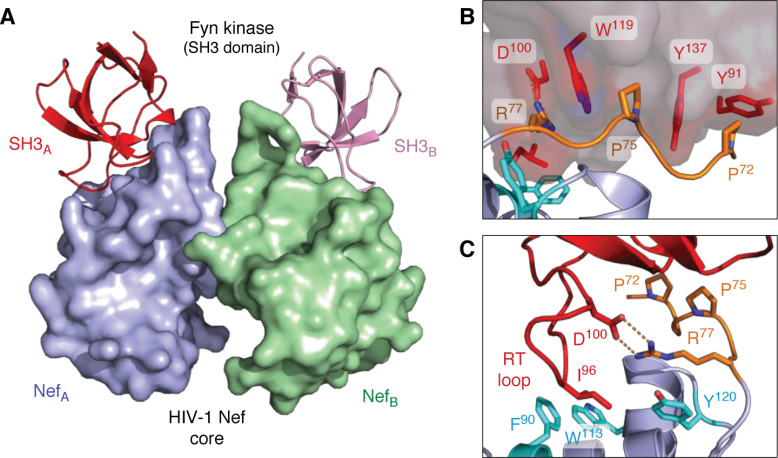
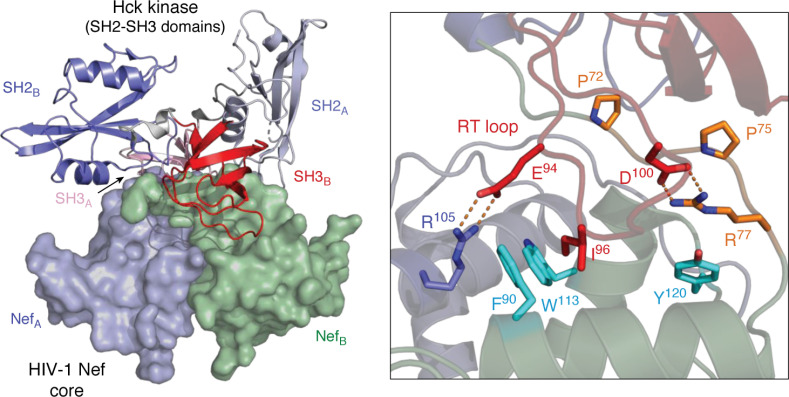
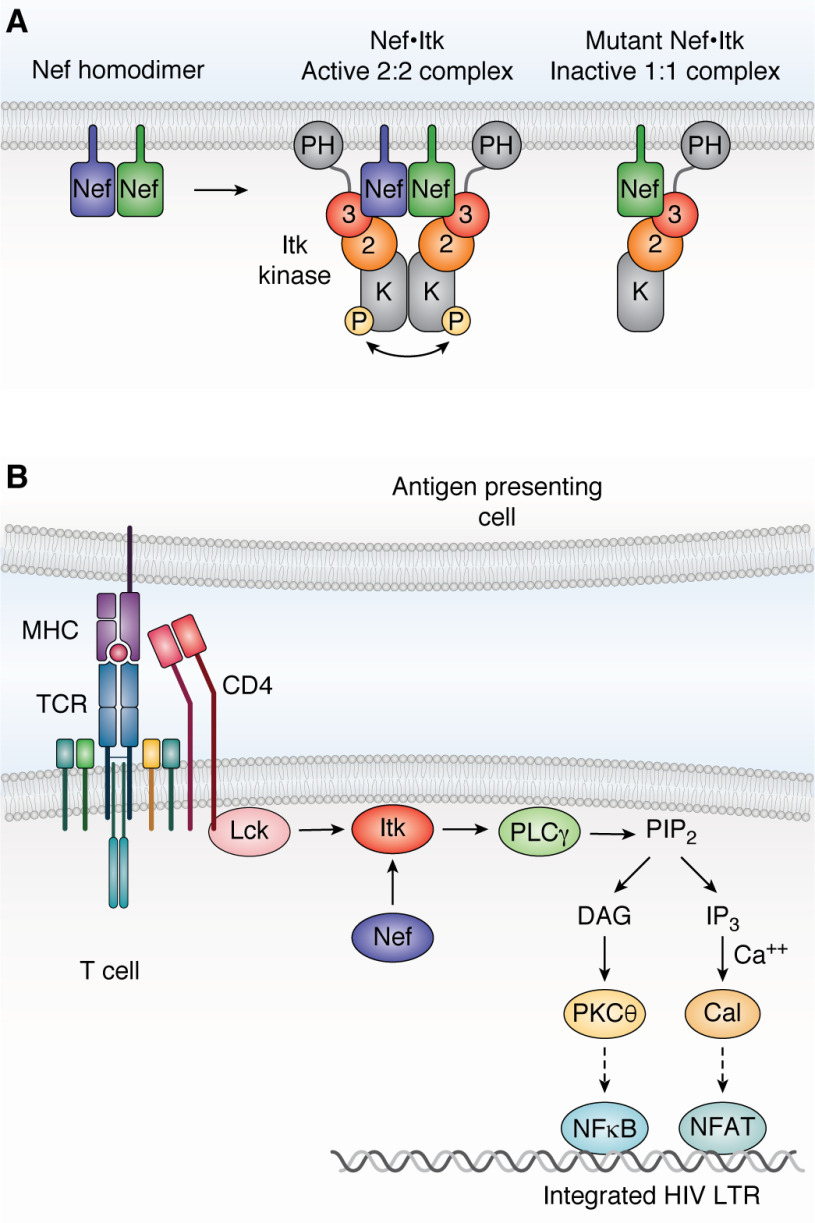
Antiretroviral therapy has revolutionized the treatment of AIDS, turning a deadly disease into a manageable chronic condition. Life-long treatment is required because existing drugs do not eradicate HIV-infected cells. The emergence of drug-resistant viral strains and uncertain vaccine prospects highlight the pressing need for new therapeutic approaches with the potential to clear the virus. The HIV-1 accessory protein Nef is essential for viral pathogenesis, making it a promising target for antiretroviral drug discovery. Nef enhances viral replication and promotes immune escape of HIV-infected cells but lacks intrinsic enzymatic activity. Instead, Nef works through diverse interactions with host cell proteins primarily related to kinase signaling pathways and endosomal trafficking. This review emphasizes the structure, function, and biological relevance of Nef interactions with host cell protein-tyrosine kinases in the broader context of Nef functions related to enhancement of the viral life cycle and immune escape. Drug discovery targeting Nef-mediated kinase activation has allowed identification of promising inhibitors of multiple Nef functions. Pharmacological inhibitors of Nef-induced MHC-I down-regulation restore the adaptive immune response to HIV-infected cells in vitro and have the potential to enhance immune recognition of latent viral reservoirs as part of a strategy for HIV clearance.
Keywords: AIDS; Btk; CD4; HIV-1 Nef; Hck; Itk; MHC-I; SH2 domain; SH3 domain; Src homology 2 domain (SH2 domain); Src homology 3 domain (SH3 domain); Src-family kinases; Tec-family kinases; bimolecular fluorescence complementation (BiFC); dimerization; endocytosis; human immunodeficiency virus (HIV); infectious disease; major histocompatibility complex (MHC); protein-protein interaction.
© 2020 Staudt et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
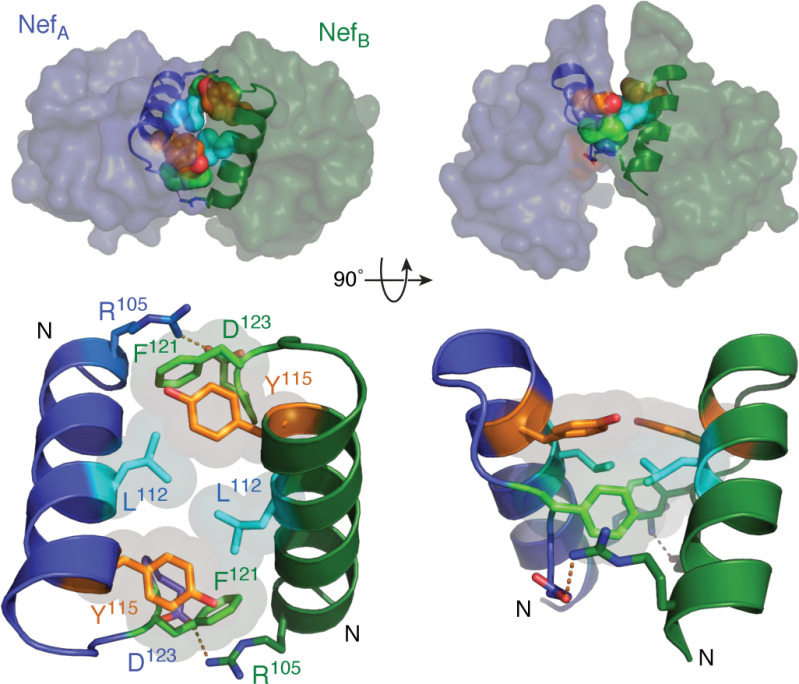
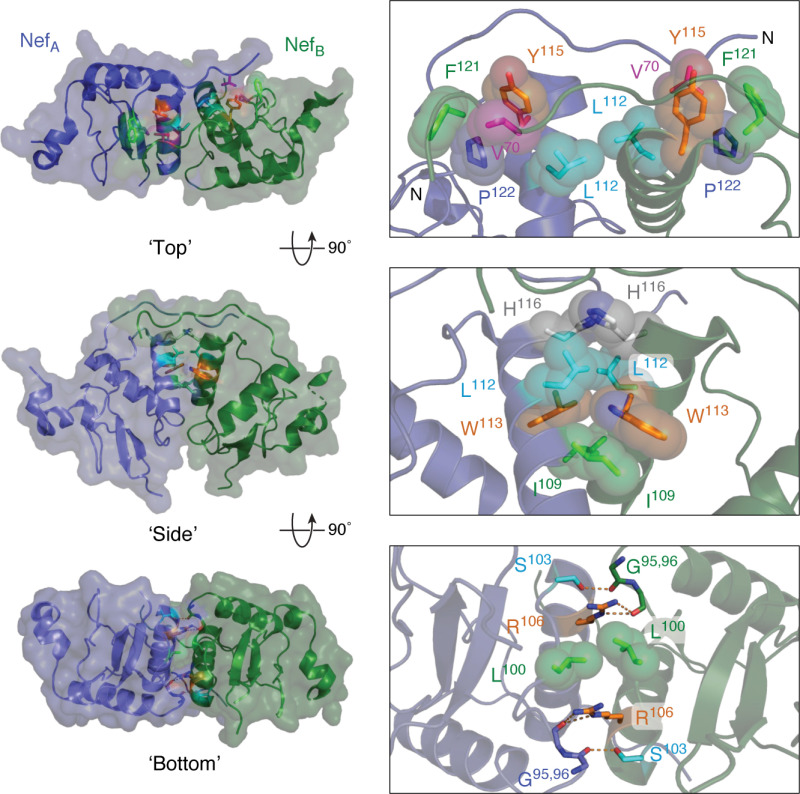
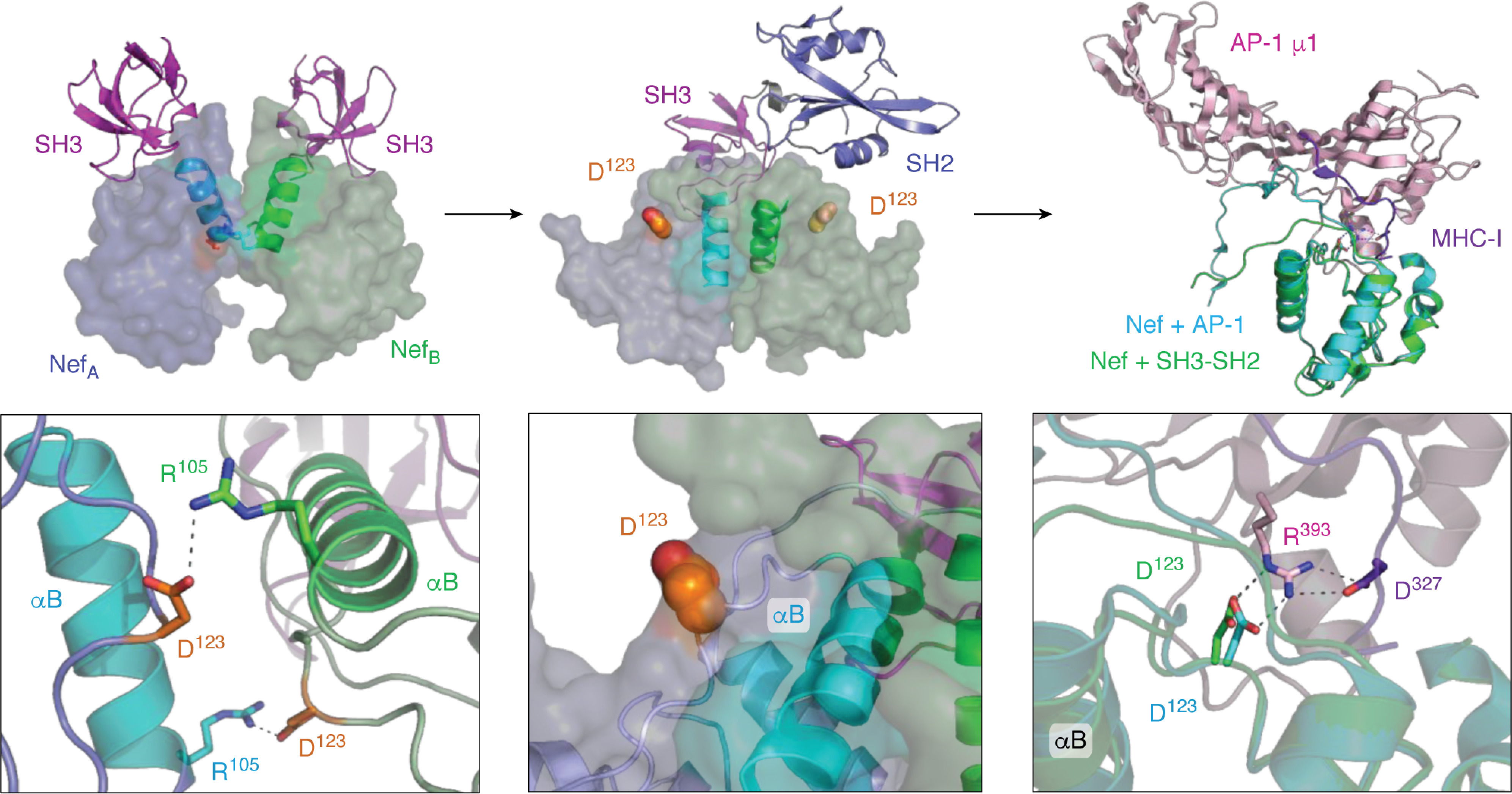
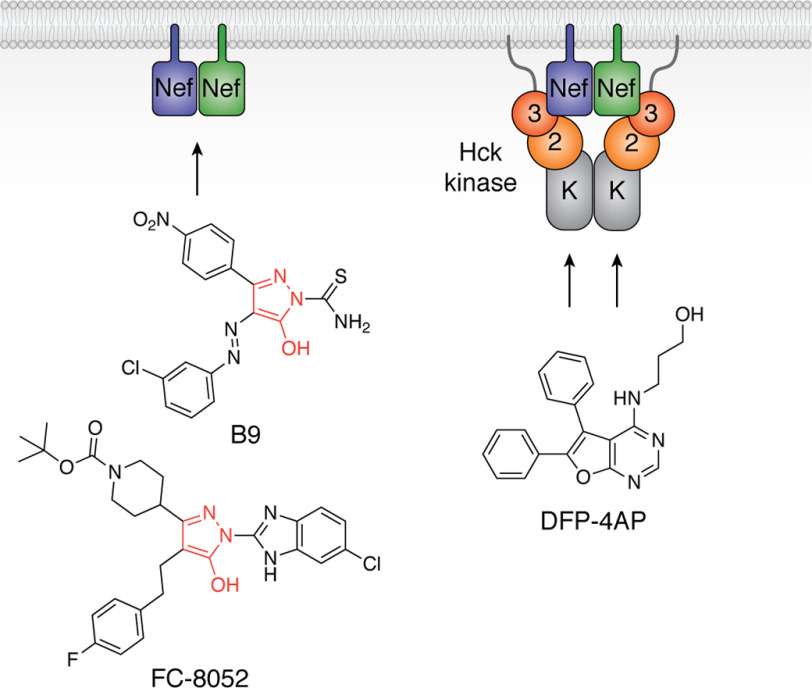
References
-
- Hammer S. M., Katzenstein D. A., Hughes M. D., Gundacker H., Schooley R. T., Haubrich R. H., Henry W. K., Lederman M. M., Phair J. P., Niu M., Hirsch M. S., and Merigan T. C. (1996) A trial comparing nucleoside monotherapy with combination therapy in HIV-infected adults with CD4 cell counts from 200 to 500 per cubic millimeter. AIDS Clinical Trials Group Study 175 Study Team. N. Engl. J. Med. 335, 1081–1090 10.1056/NEJM199610103351501 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
