

Regulation of Nrf2/Keap1 signalling in human skeletal muscle during exercise to exhaustion in normoxia, severe acute hypoxia and post-exercise ischaemia: Influence of metabolite accumulation and oxygenation
- PMID: 32863217
- PMCID: PMC7358388
- DOI: 10.1016/j.redox.2020.101627
Regulation of Nrf2/Keap1 signalling in human skeletal muscle during exercise to exhaustion in normoxia, severe acute hypoxia and post-exercise ischaemia: Influence of metabolite accumulation and oxygenation
Abstract
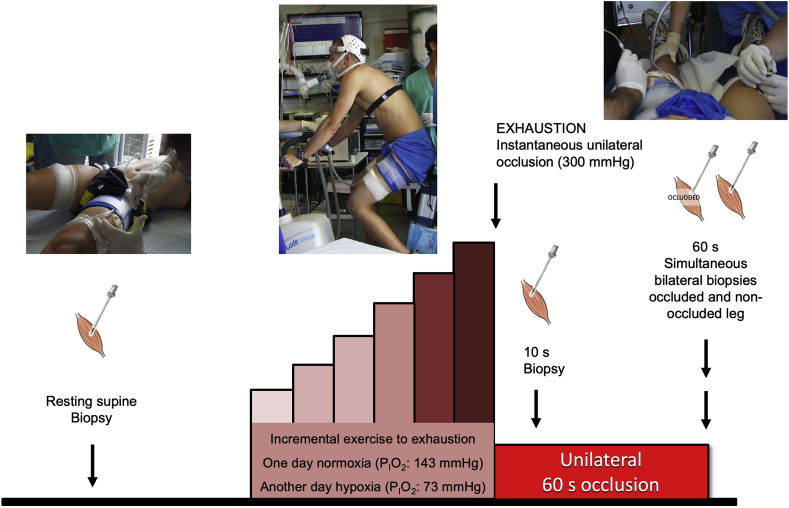
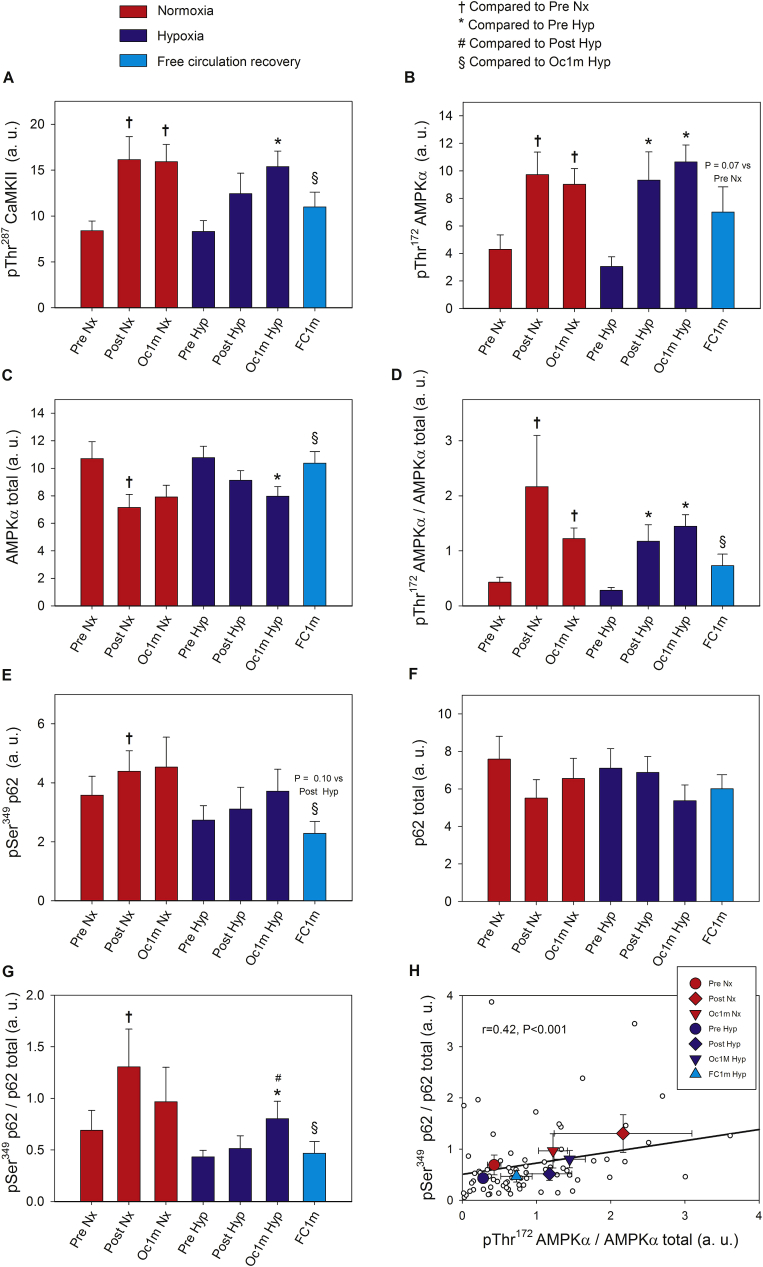
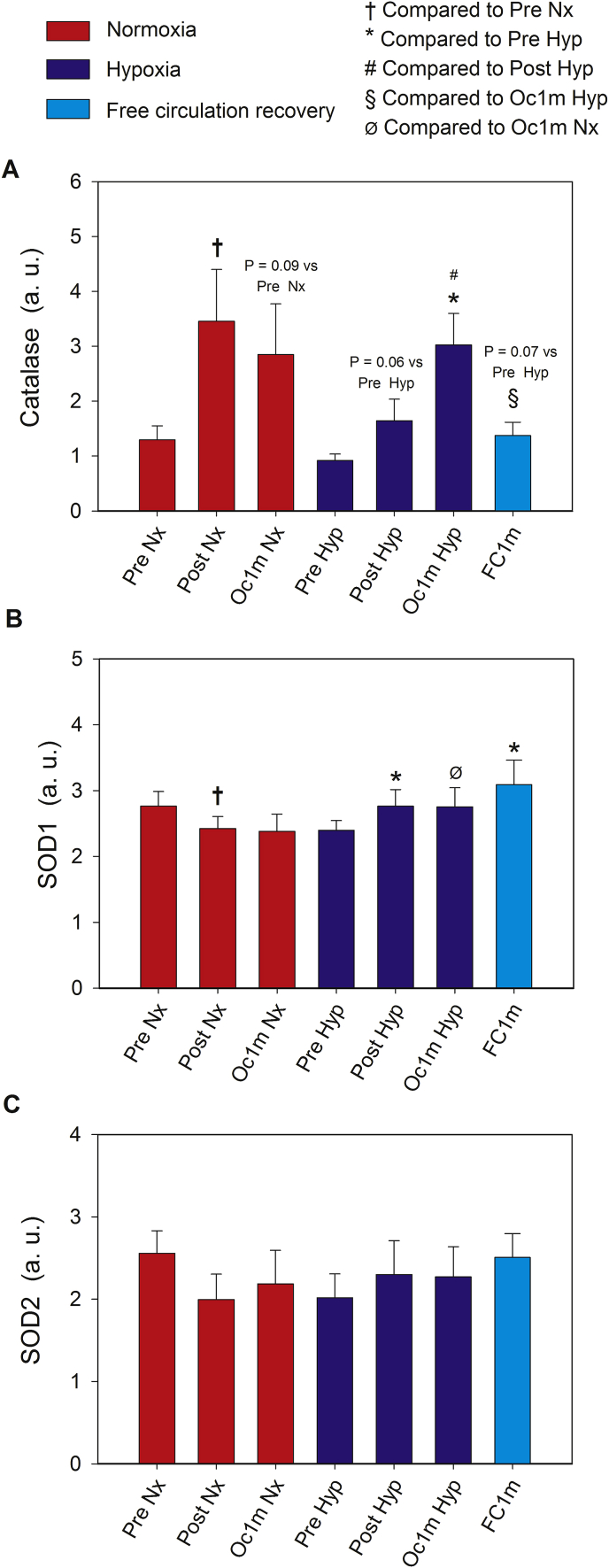
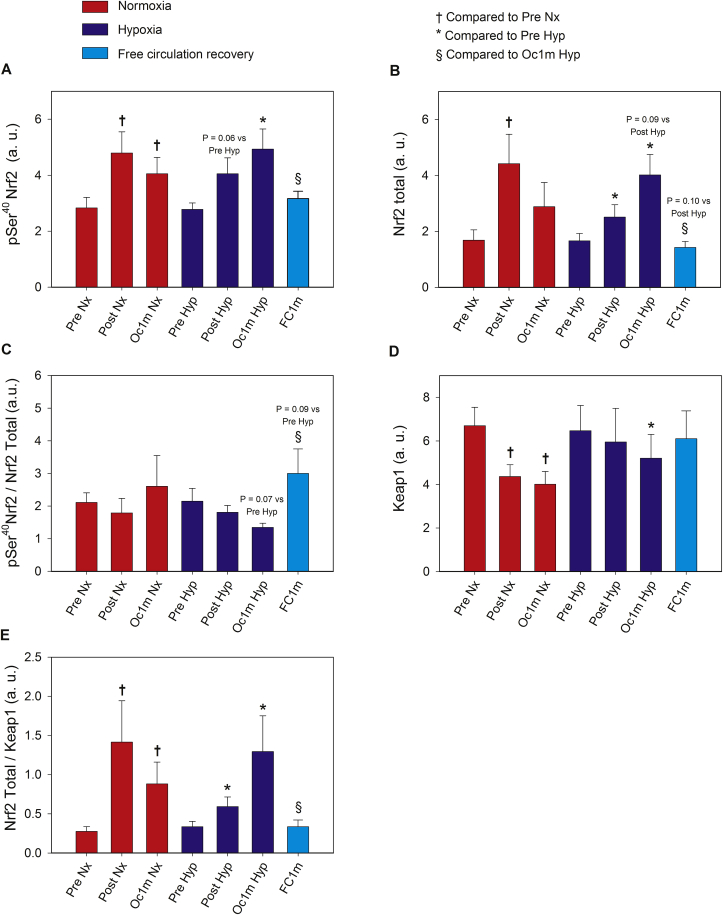
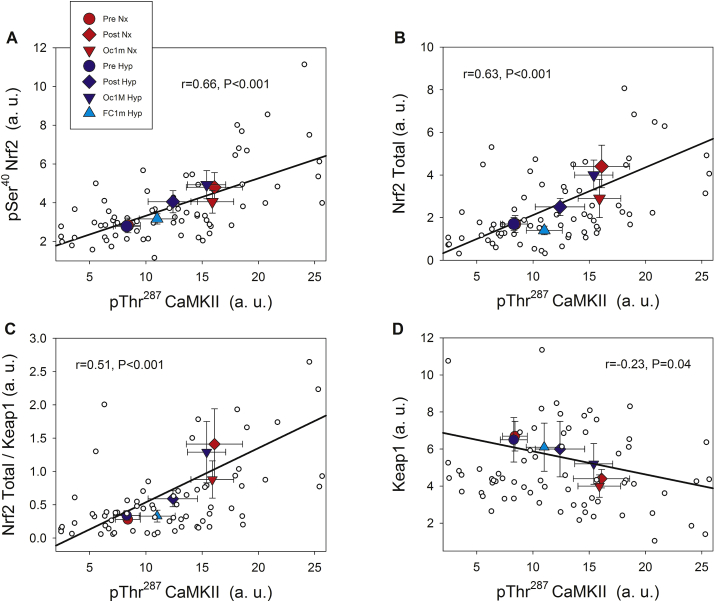
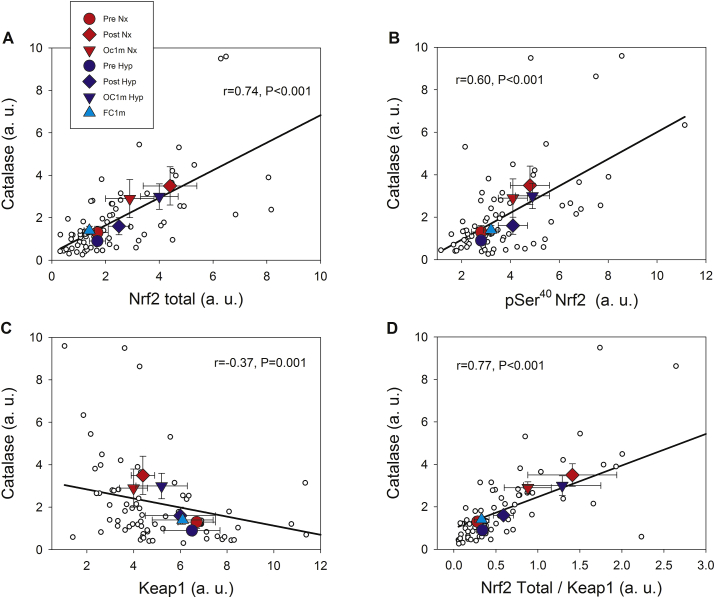
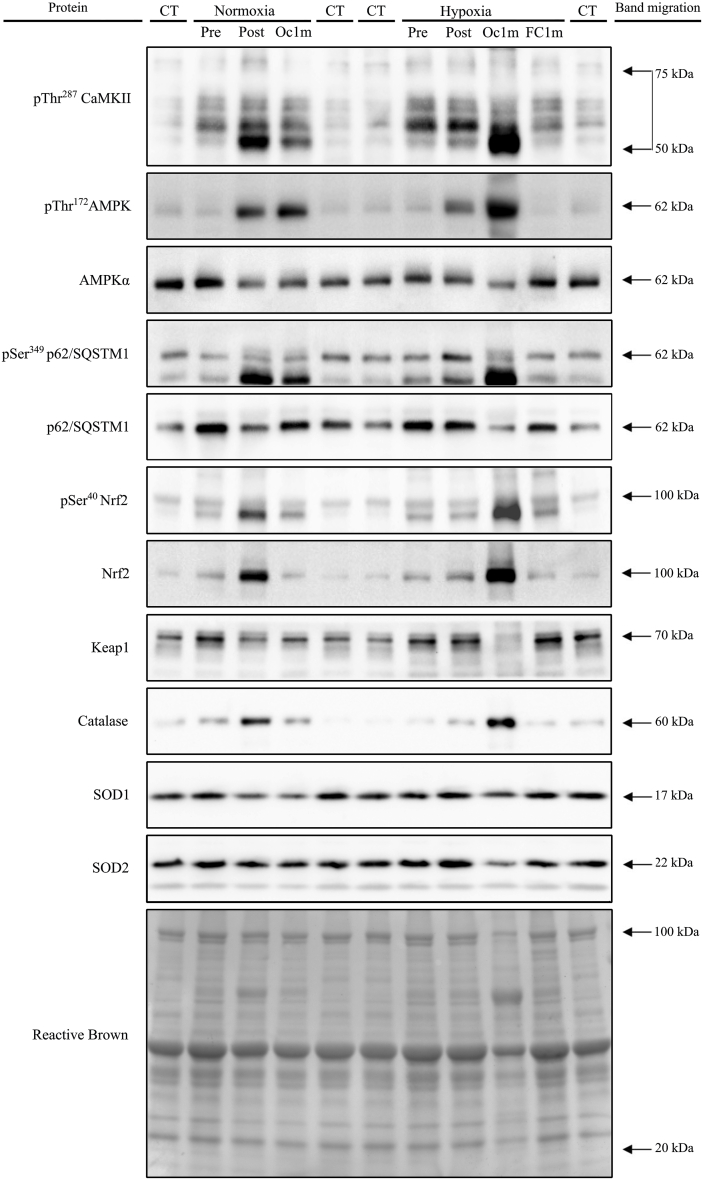
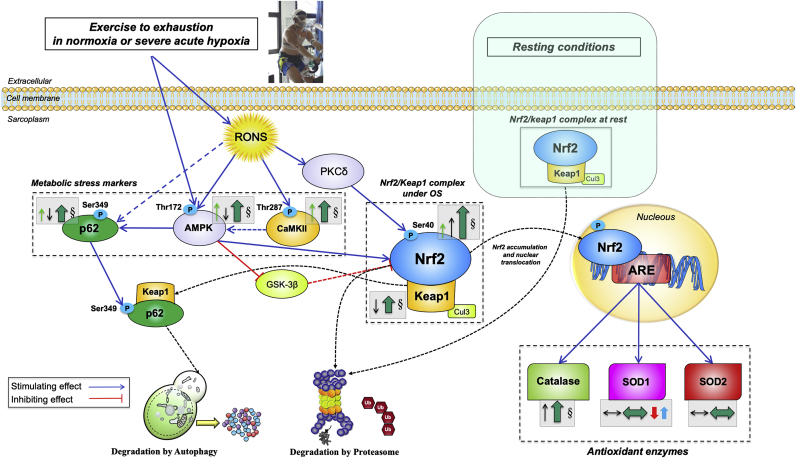
The Nrf2 transcription factor is induced by reactive oxygen and nitrogen species and is necessary for the adaptive response to exercise in mice. It remains unknown whether Nrf2 signalling is activated by exercise in human skeletal muscle. Here we show that Nrf2 signalling is activated by exercise to exhaustion with similar responses in normoxia (PIO2: 143 mmHg) and severe acute hypoxia (PIO2: 73 mmHg). CaMKII and AMPKα phosphorylation were similarly induced in both conditions. Enhanced Nrf2 signalling was achieved by raising Nrf2 total protein and Ser40 Nrf2 phosphorylation, accompanied by a reduction of Keap1. Keap1 protein degradation is facilitated by the phosphorylation of p62/SQSTM1 at Ser349 by AMPK, which targets Keap1 for autophagic degradation. Consequently, the Nrf2-to-Keap1 ratio was markedly elevated and closely associated with a 2-3-fold increase in Catalase protein. No relationship was observed between Nrf2 signalling and SOD1 and SOD2 protein levels. Application of ischaemia immediately at the end of exercise maintained these changes, which were reverted within 1 min of recovery with free circulation. While SOD2 did not change significantly during either exercise or ischaemia, SOD1 protein expression was marginally downregulated and upregulated during exercise in normoxia and hypoxia, respectively. We conclude that Nrf2/Keap1/Catalase pathway is rapidly regulated during exercise and recovery in human skeletal muscle. Catalase emerges as an essential antioxidant enzyme acutely upregulated during exercise and ischaemia. Post-exercise ischaemia maintains Nrf2 signalling at the level reached at exhaustion and can be used to avoid early post-exercise recovery, which is O2-dependent.
Keywords: AMPK; CaMKII; Fatigue; High-intensity exercise; Ischaemia; Performance.
Copyright © 2020 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Morales-Alamo D., Calbet J.A. Free radicals and sprint exercise in humans. Free Radic. Res. 2014;48(1):30–42. - PubMed
-
- Morales-Alamo D., Ponce-Gonzalez J.G., Guadalupe-Grau A., Rodriguez-Garcia L., Santana A., Cusso R. Critical role for free radicals on sprint exercise-induced CaMKII and AMPKalpha phosphorylation in human skeletal muscle. J. Appl. Physiol. 2013;114(5):566–577. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
