

Melatonin sensitises shikonin-induced cancer cell death mediated by oxidative stress via inhibition of the SIRT3/SOD2-AKT pathway
- PMID: 32863233
- PMCID: PMC7358455
- DOI: 10.1016/j.redox.2020.101632
Melatonin sensitises shikonin-induced cancer cell death mediated by oxidative stress via inhibition of the SIRT3/SOD2-AKT pathway
Abstract
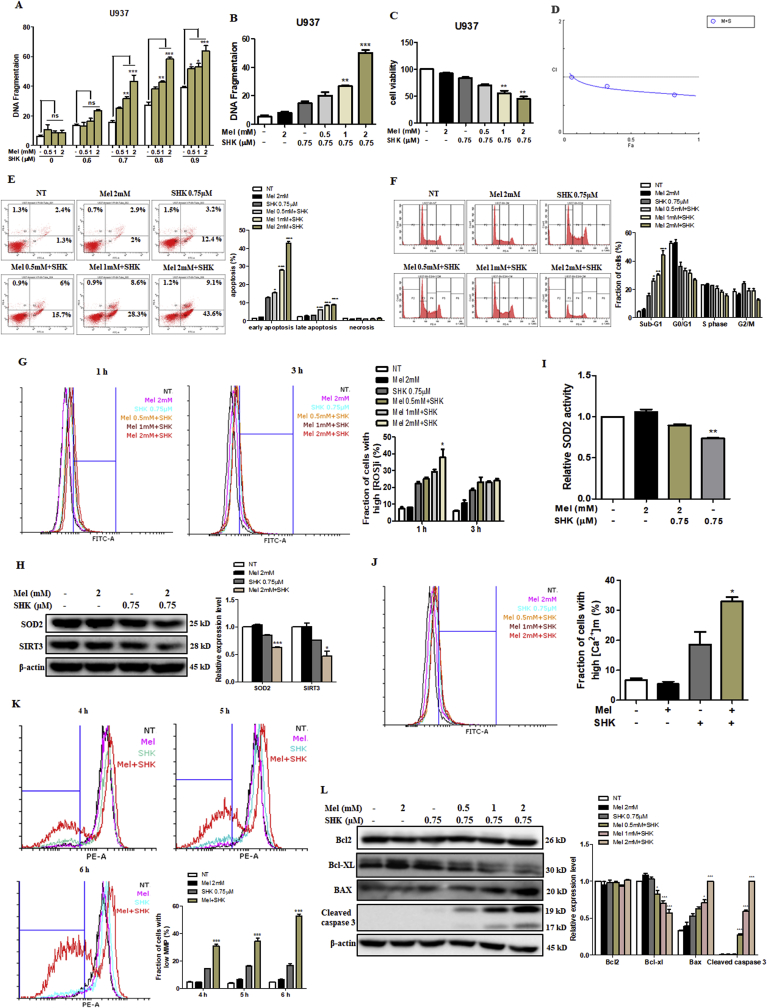
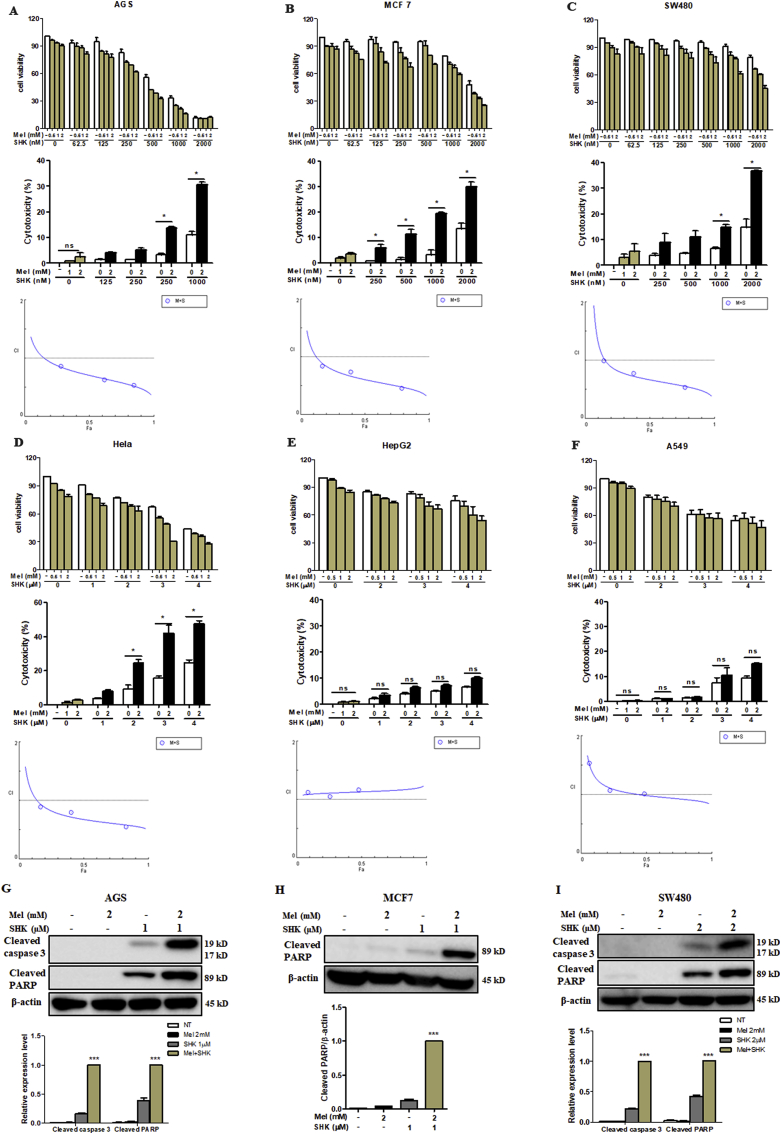
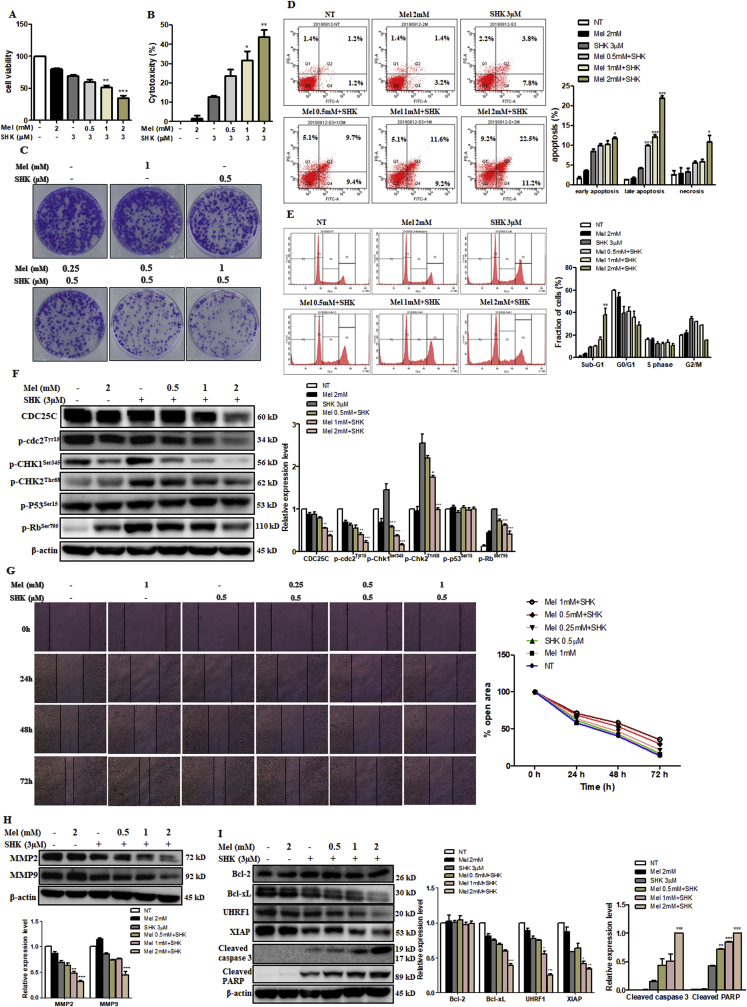
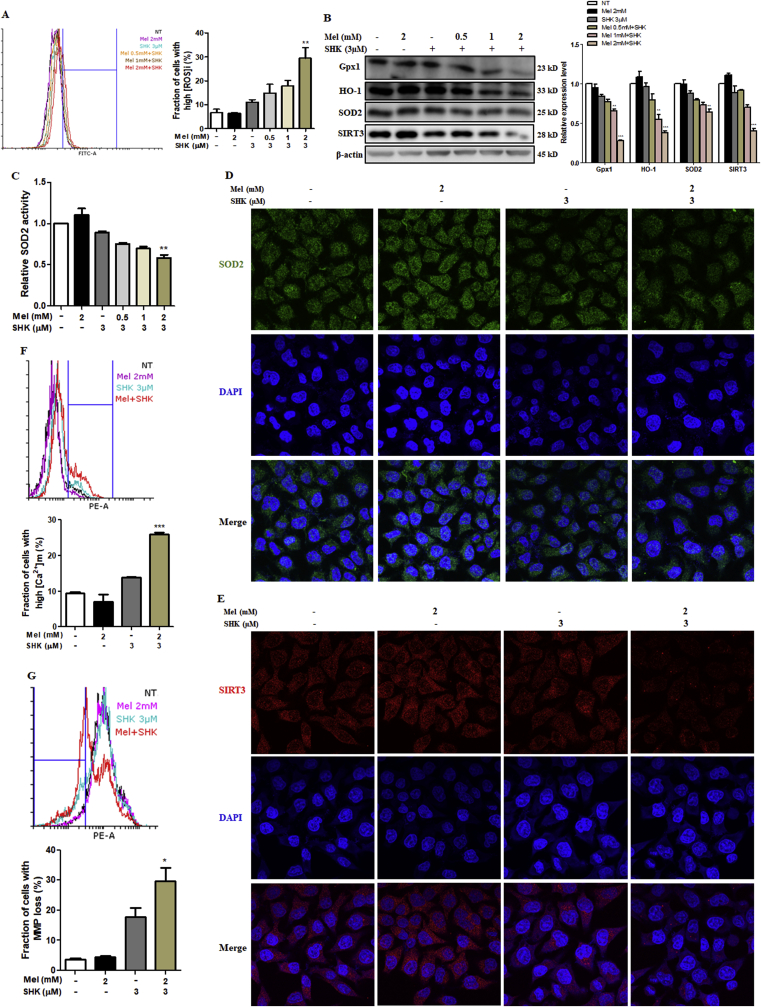
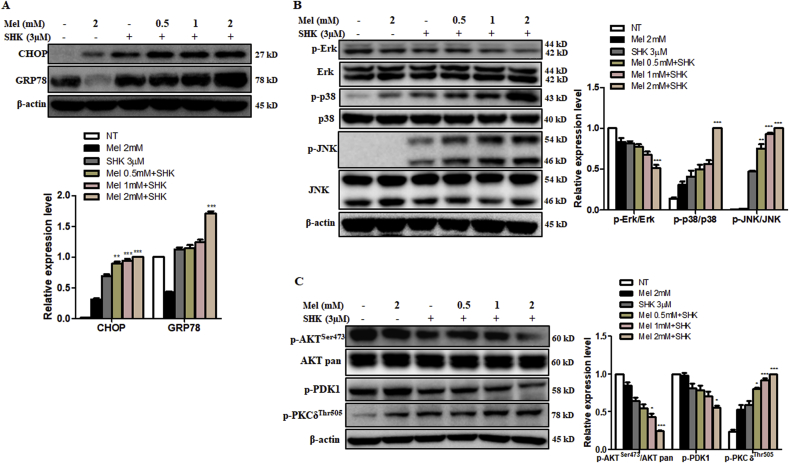
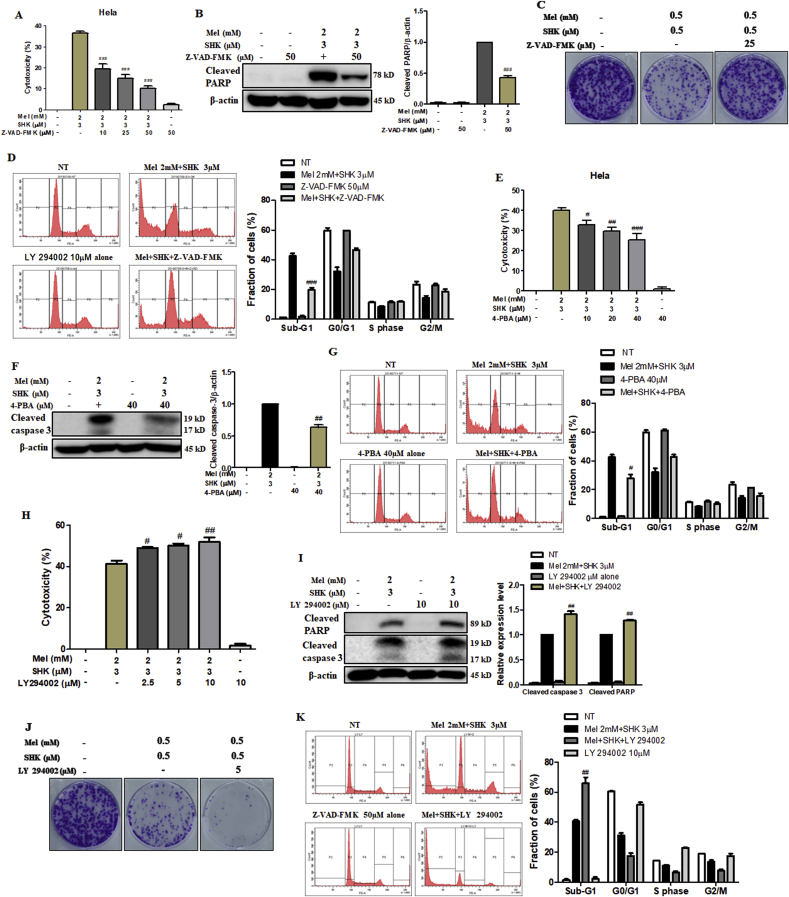
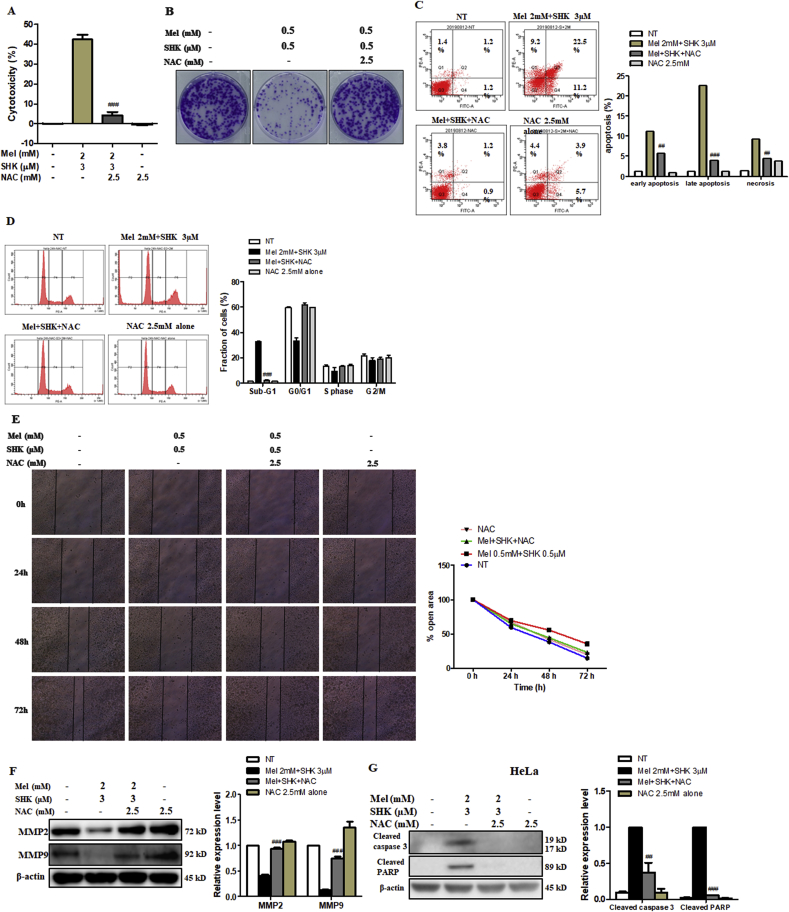
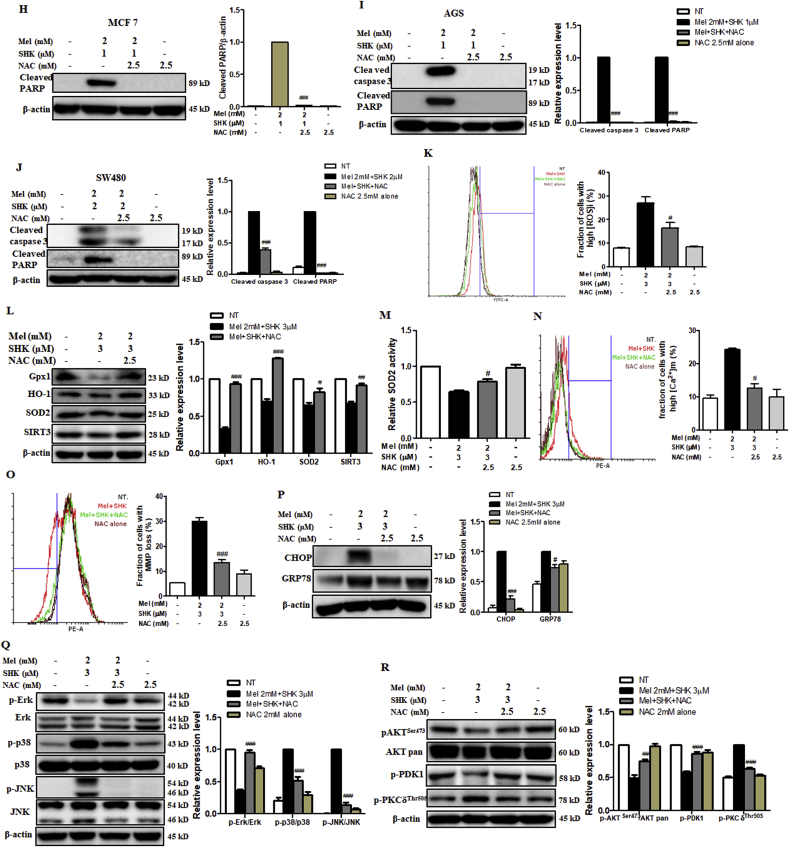
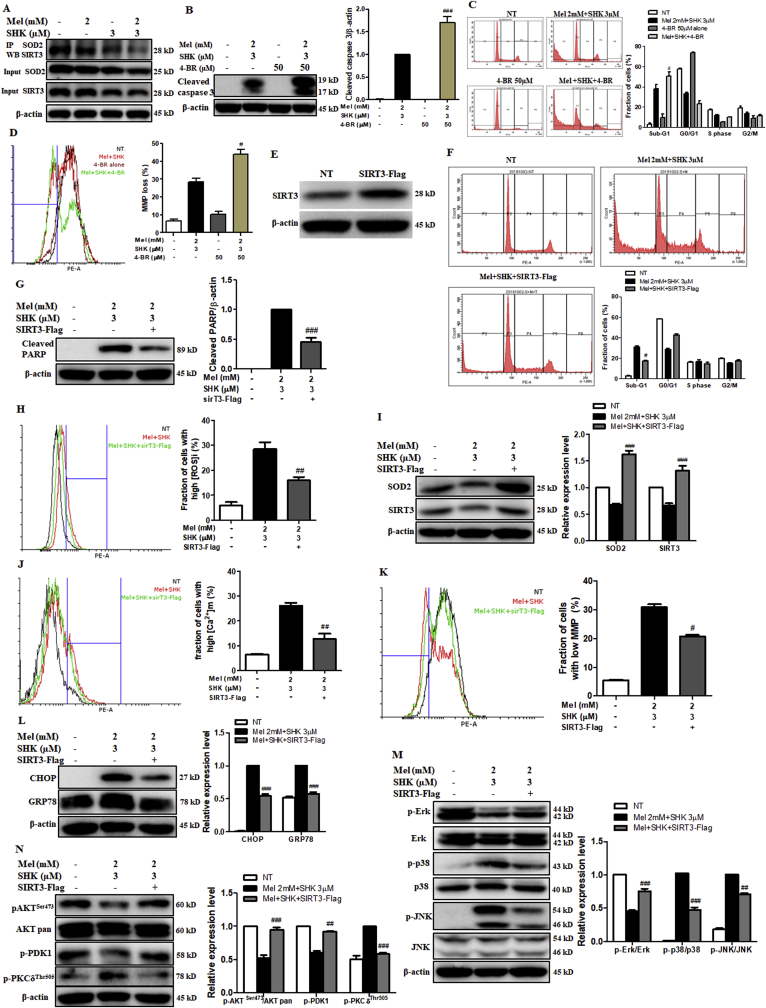
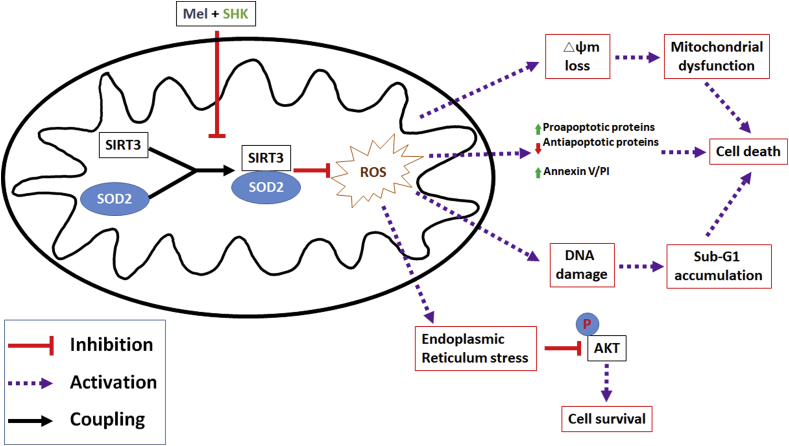
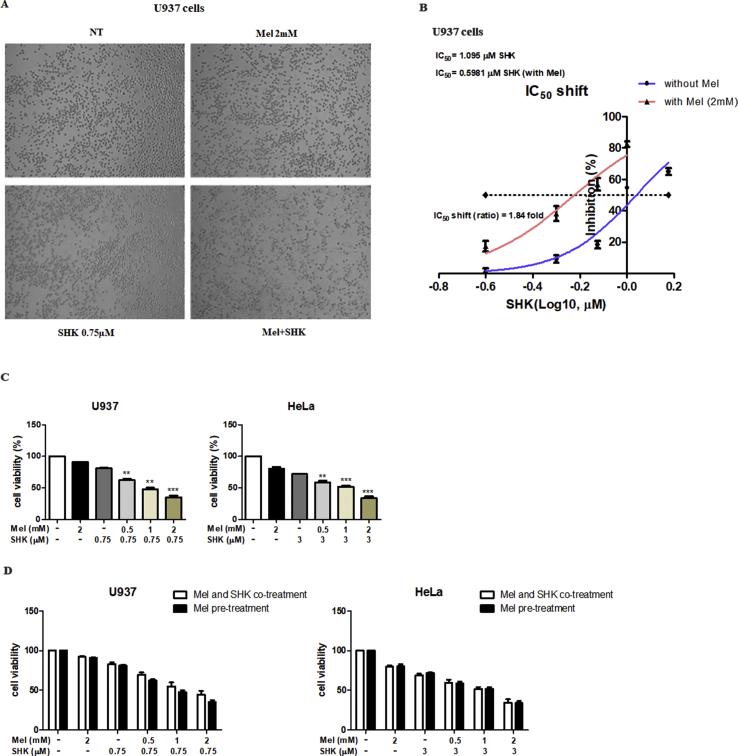
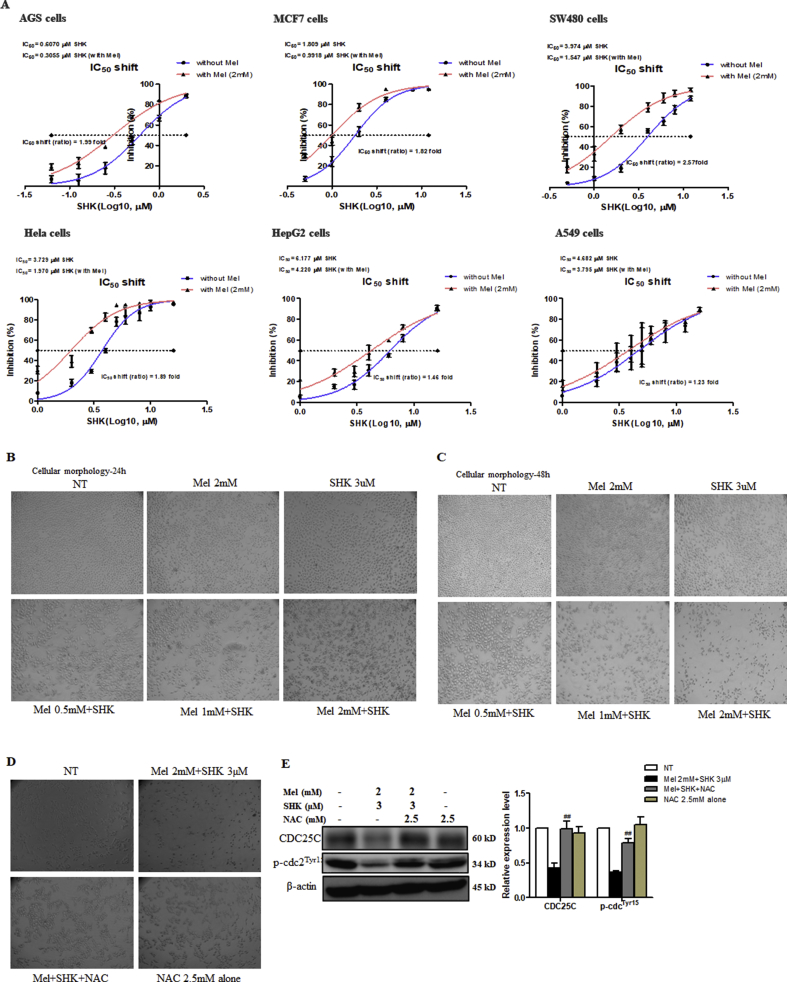
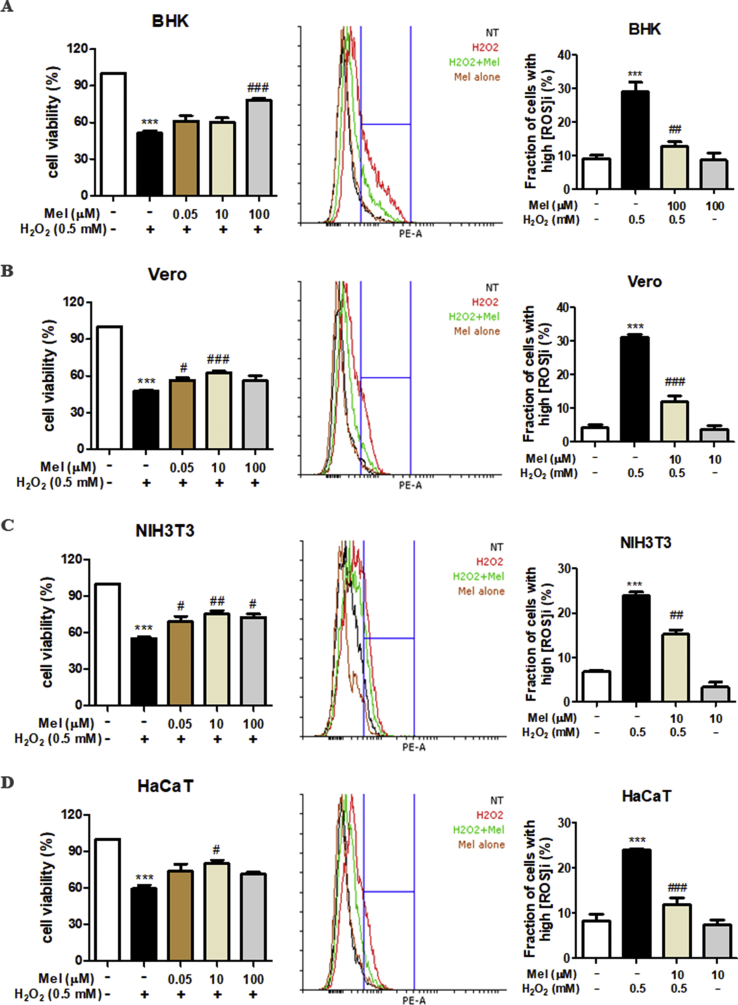
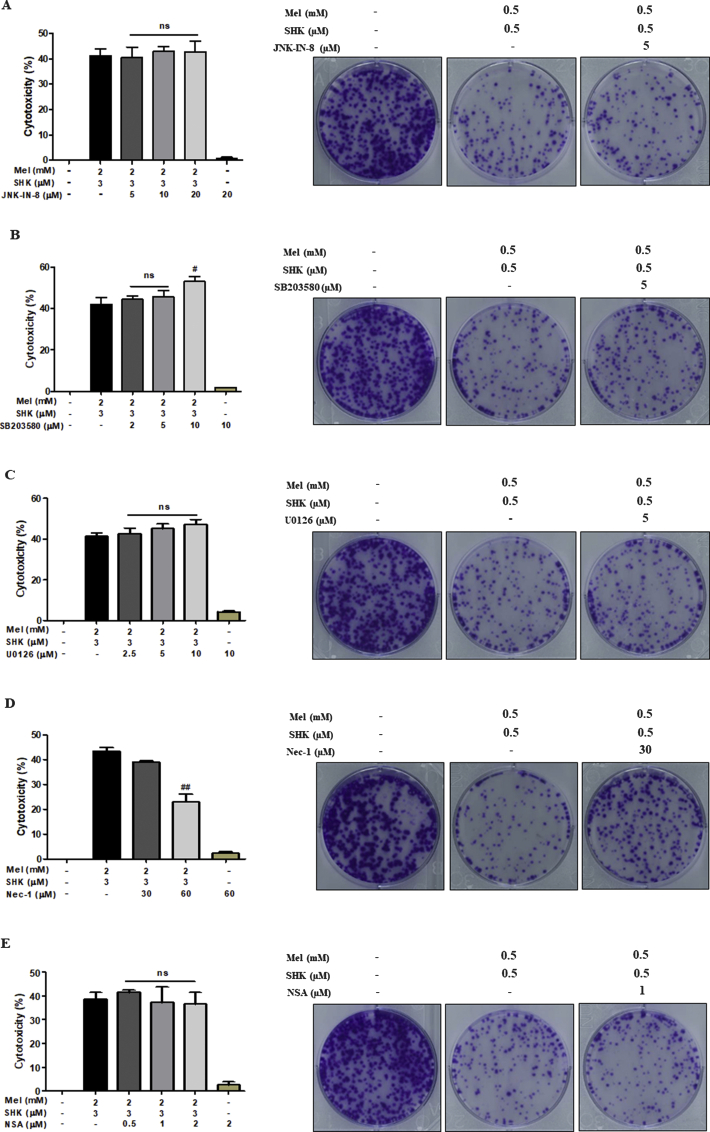
Recent research suggests that melatonin (Mel), an endogenous hormone and natural supplement, possesses anti-proliferative effects and can sensitise cells to anti-cancer therapies. Although shikonin (SHK) also possesses potential anti-cancer properties, the poor solubility and severe systemic toxicity of this compound hinders its clinical usage. In this study, we combined Mel and SHK, a potentially promising chemotherapeutic drug combination, with the aim of reducing the toxicity of SHK and enhancing the overall anti-cancer effects. We demonstrate for the first time that Mel potentiates the cytotoxic effects of SHK on cancer cells by inducing oxidative stress via inhibition of the SIRT3/SOD2-AKT pathway. Particularly, Mel-SHK treatment induced oxidative stress, increased mitochondrial calcium accumulation and reduced the mitochondrial membrane potential in various cancer cells, leading to apoptosis. This drug combination also promoted endoplasmic reticulum (ER) stress, leading to AKT dephosphorylation. In HeLa cells, Mel-SHK treatment reduced SIRT3/SOD2 expression and SOD2 activity, while SIRT3 overexpression dramatically reduced Mel-SHK-induced oxidative stress, ER stress, mitochondrial dysfunction and apoptosis. Hence, we propose the combination of Mel and SHK as a novel candidate chemotherapeutic regimen that targets the SIRT3/SOD2-AKT pathway in cancer.
Keywords: AKT; Apoptosis; Melatonin; Reactive oxygen species; SIRT3/SOD2; Shikonin.
Copyright © 2020 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bray F., Ferlay J., Soerjomataram I., Siegel R.L., Torre L.A., Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca - Cancer J. Clin. 2018;68(6):394–424. - PubMed
-
- Zarour L.R., Anand S., Billingsley K.G., Bisson W.H., Cercek A., Clarke M.F., Coussens L.M., Gast C.E., Geltzeiler C.B., Hansen L., Kelley K.A., Lopez C.D., Rana S.R., Ruhl R., Tsikitis V.L., Vaccaro G.M., Wong M.H., Mayo S.C. Colorectal cancer liver metastasis: evolving paradigms and future directions. Cell Mol. Gastroenterol. Hepatol. 2017;3(2):163–173. - PMC - PubMed
-
- Reiter R.J., Tan D.X. Mitochondria: the birth place, battle ground and the site of melatonin metabolism in cells. Melatonin Res. 2019;2(1):44–66.
-
- Mortezaee K., Najafi M., Farhood B., Ahmadi A., Potes Y., Shabeeb D., Musa A.E. Modulation of apoptosis by melatonin for improving cancer treatment efficiency: an updated review. Life Sci. 2019;228:228–241. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
