

Neurotoxic microglia promote TDP-43 proteinopathy in progranulin deficiency
- PMID: 32866962
- PMCID: PMC7746606
- DOI: 10.1038/s41586-020-2709-7
Neurotoxic microglia promote TDP-43 proteinopathy in progranulin deficiency
Abstract
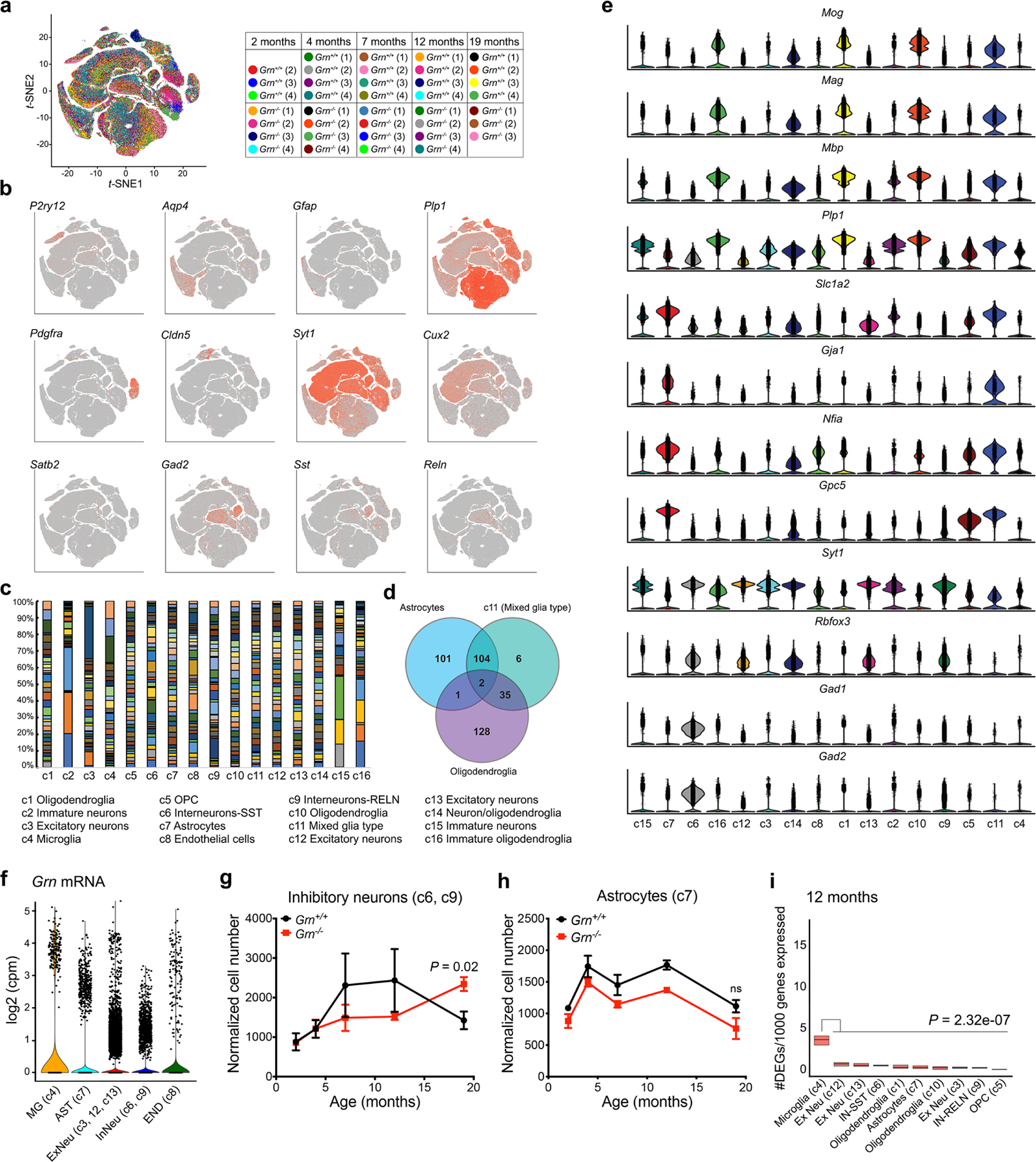
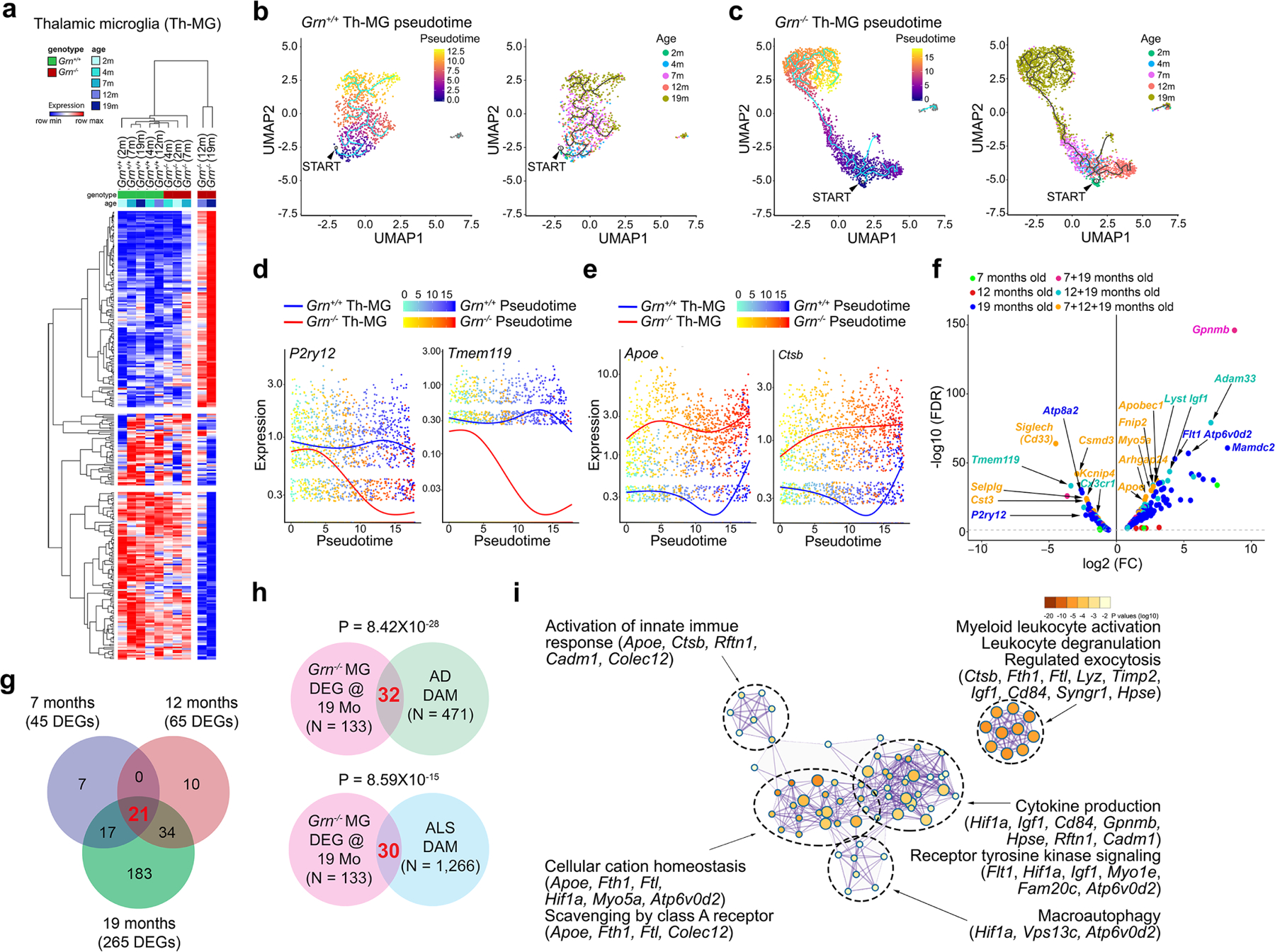
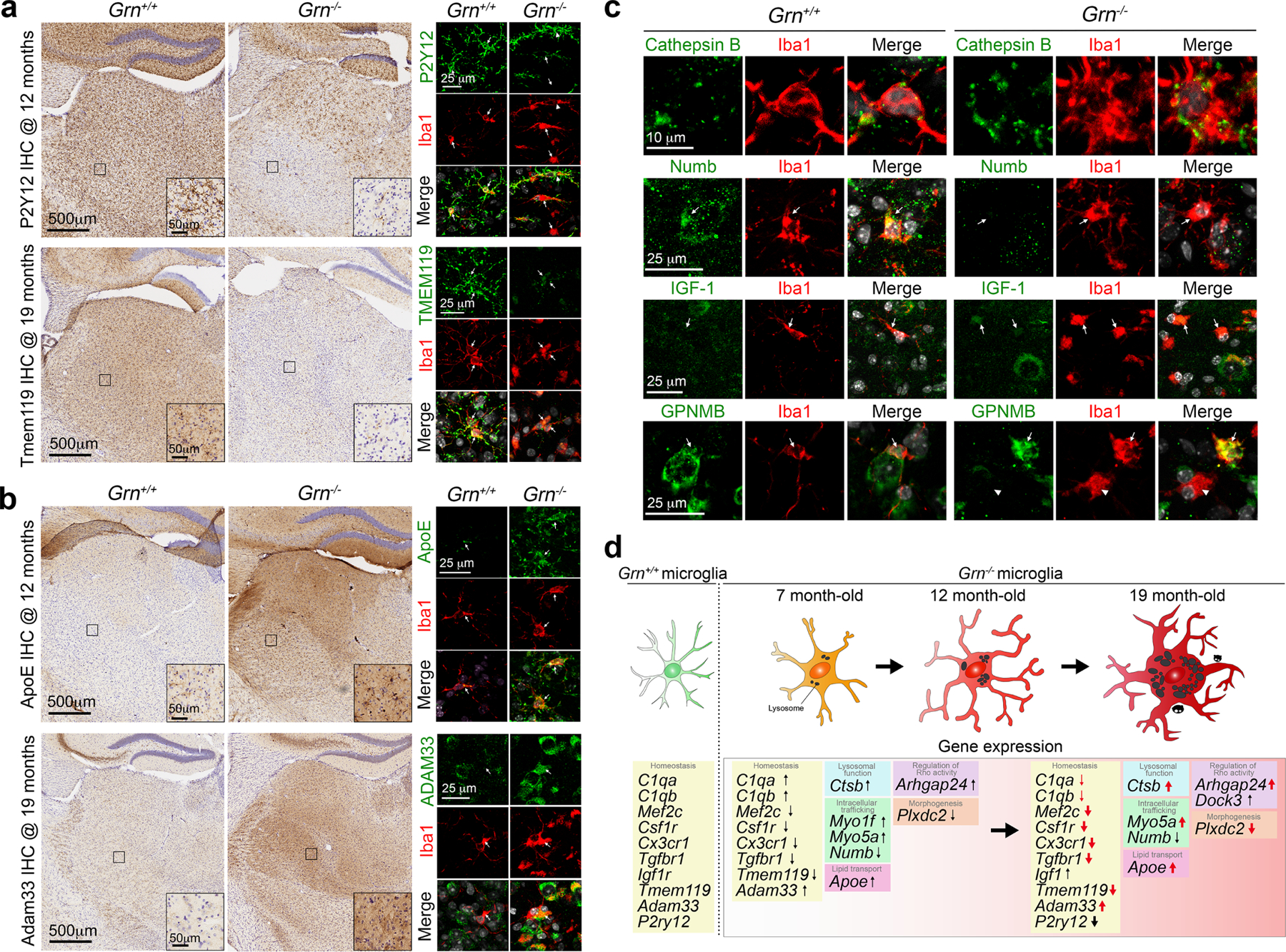
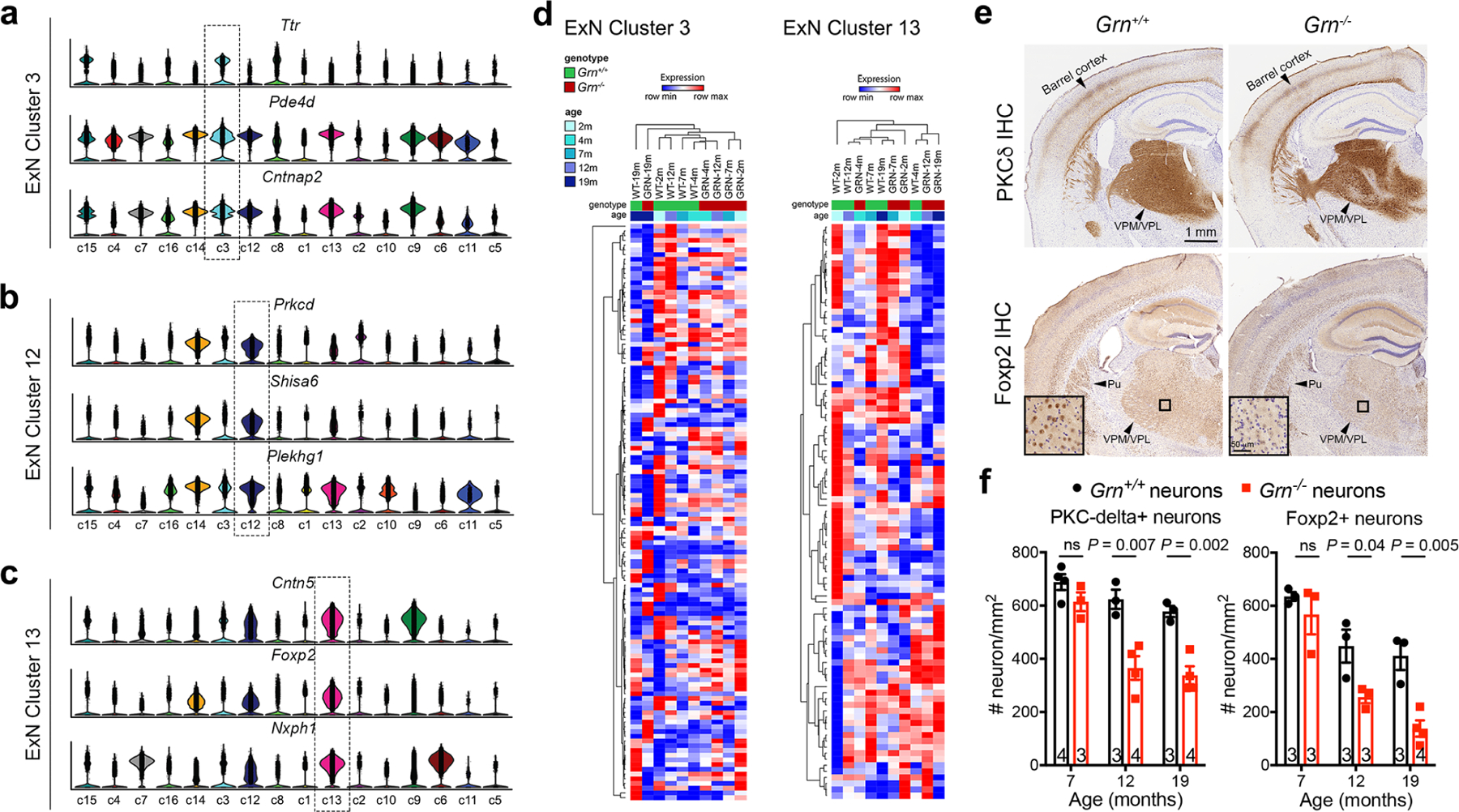
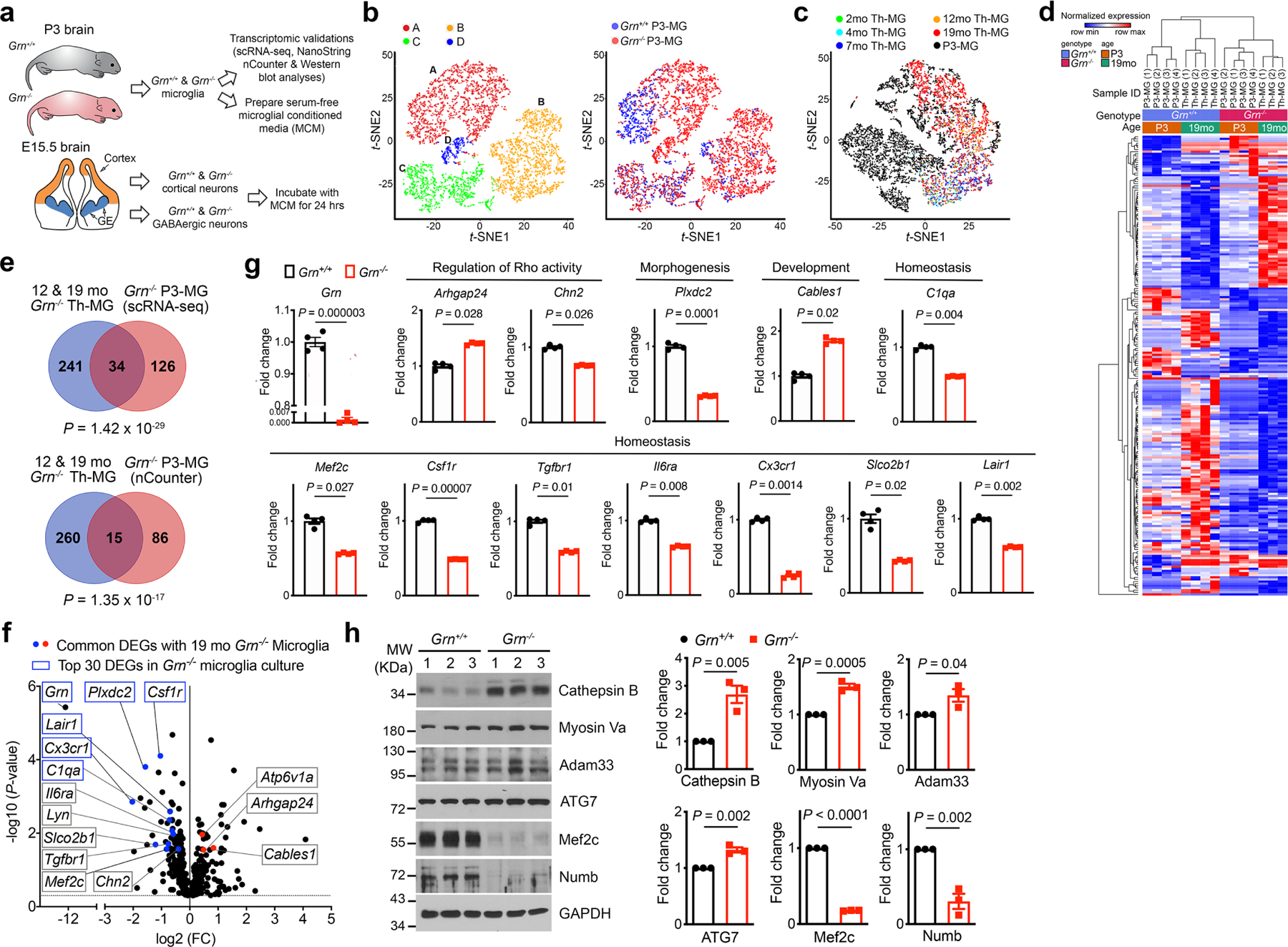
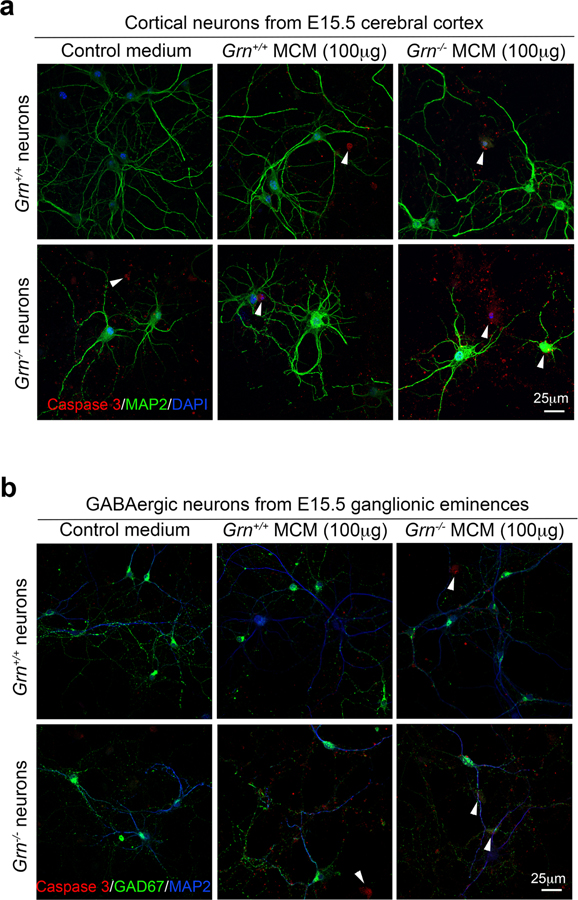
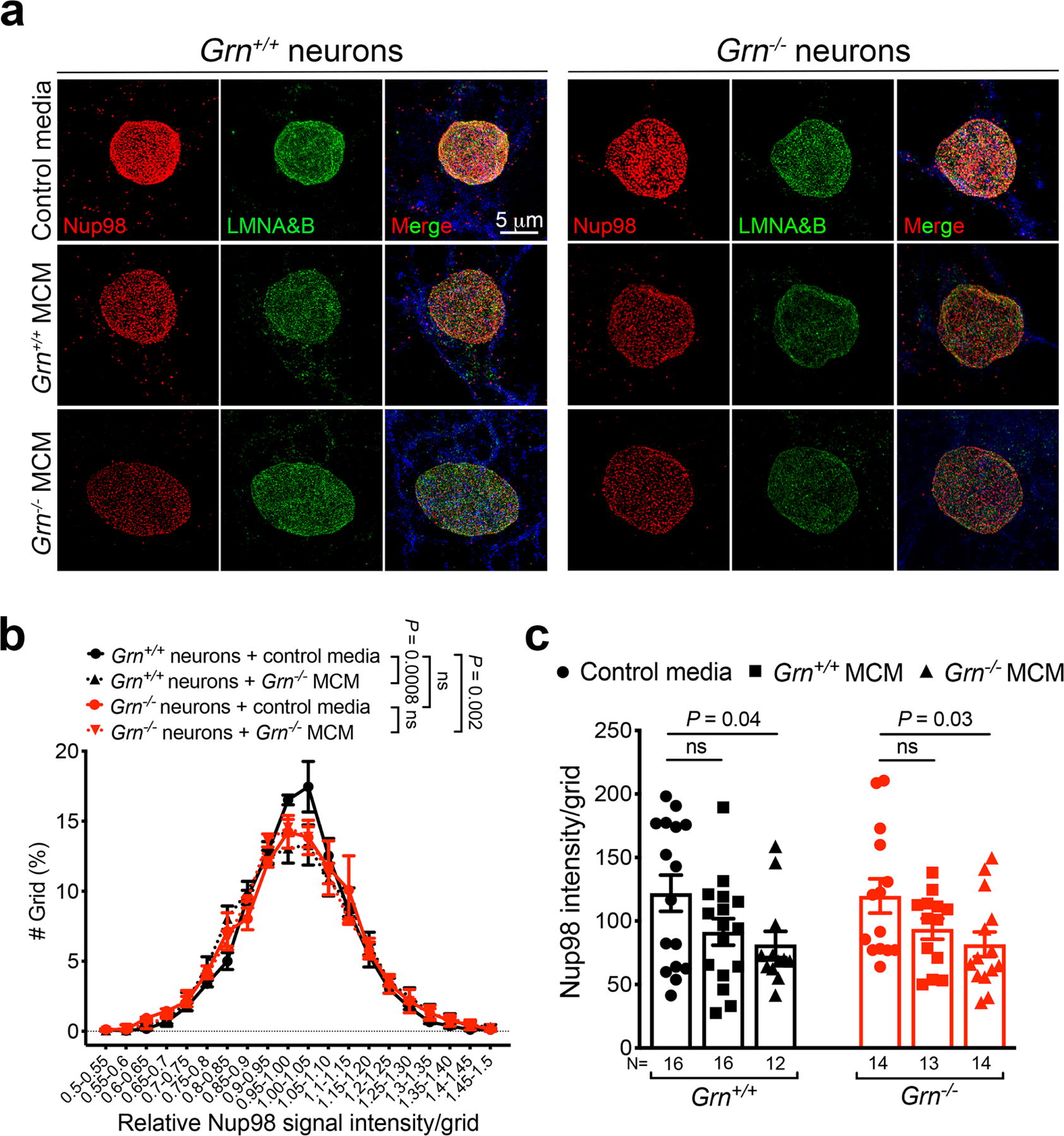
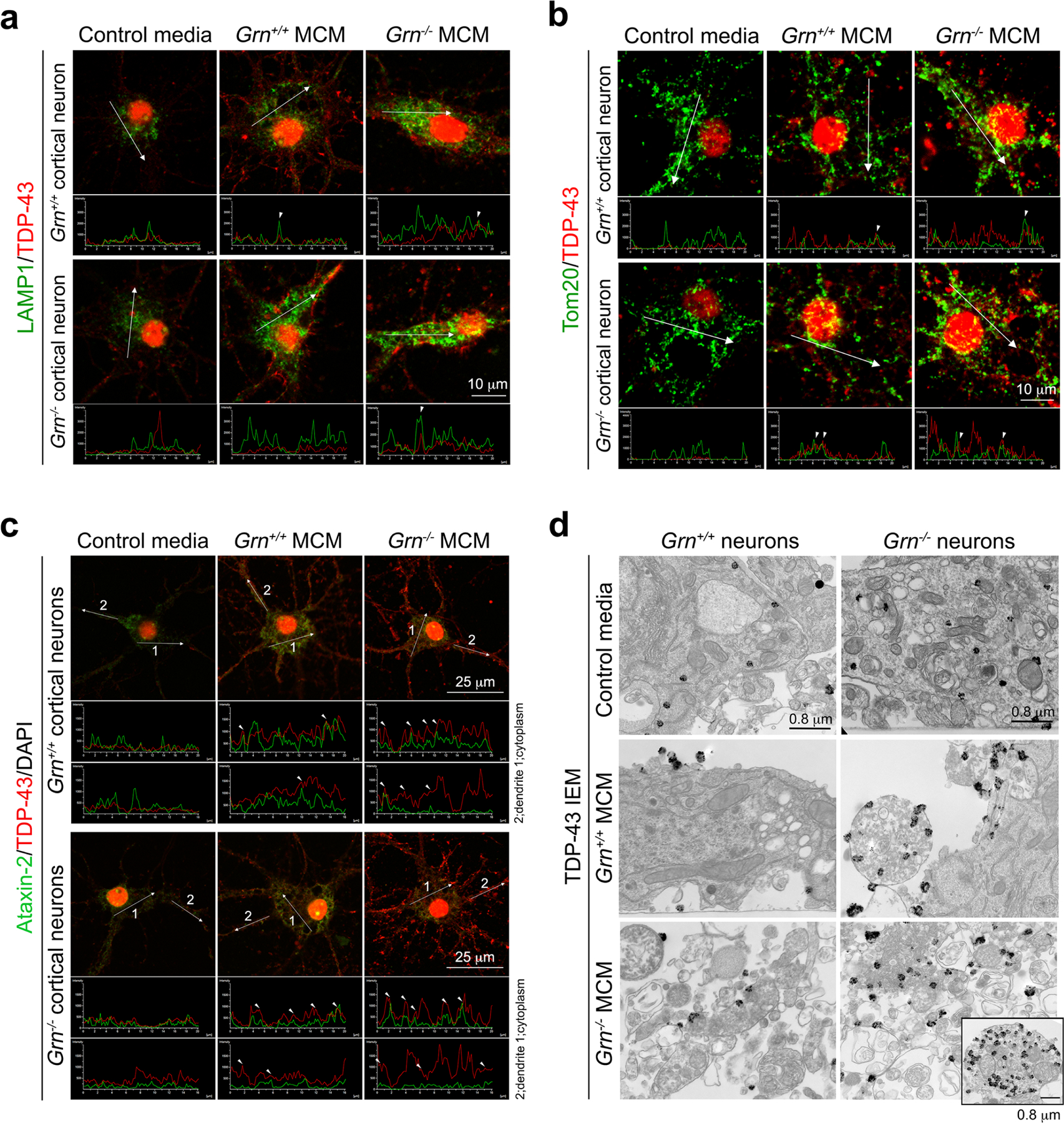
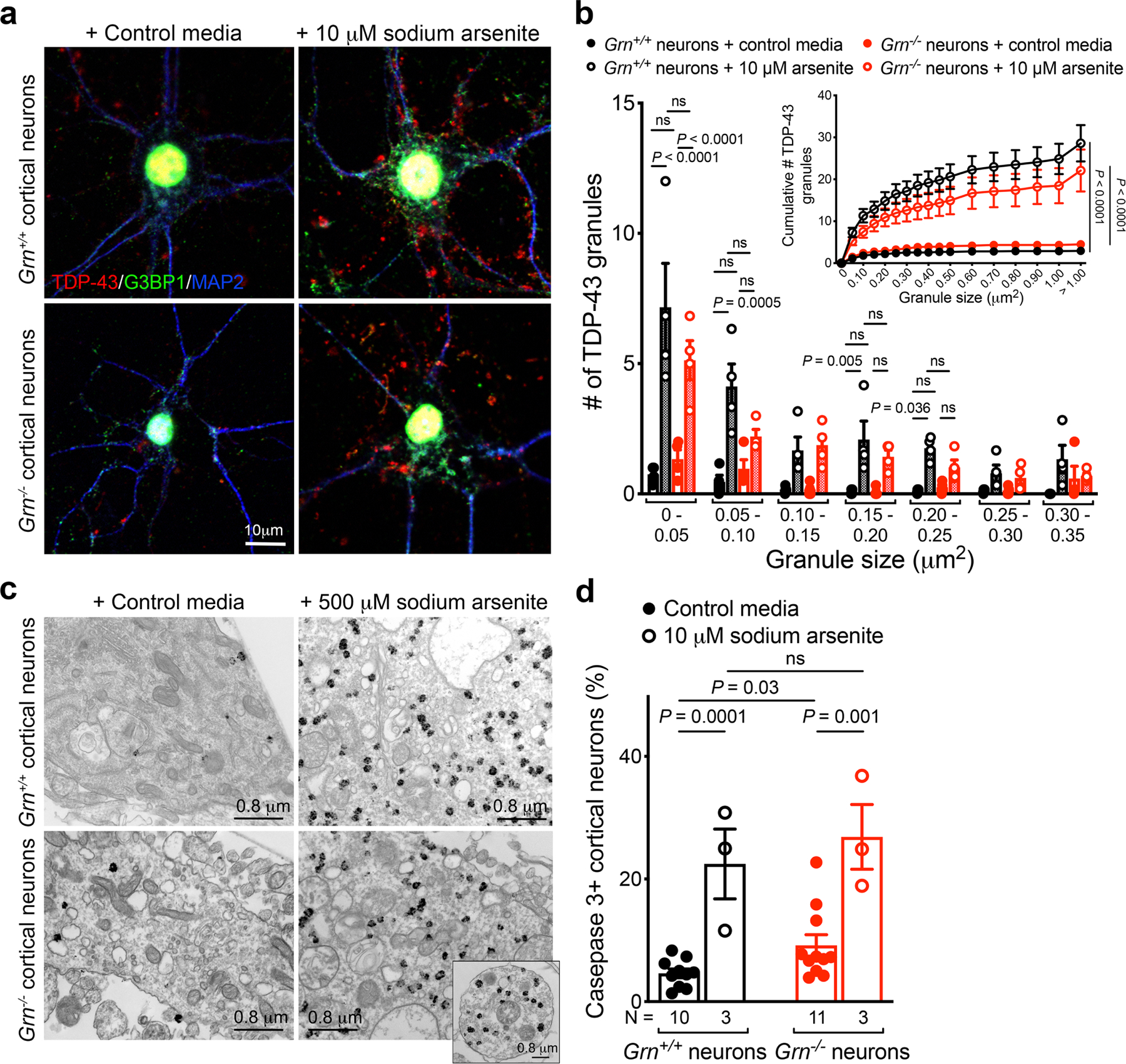
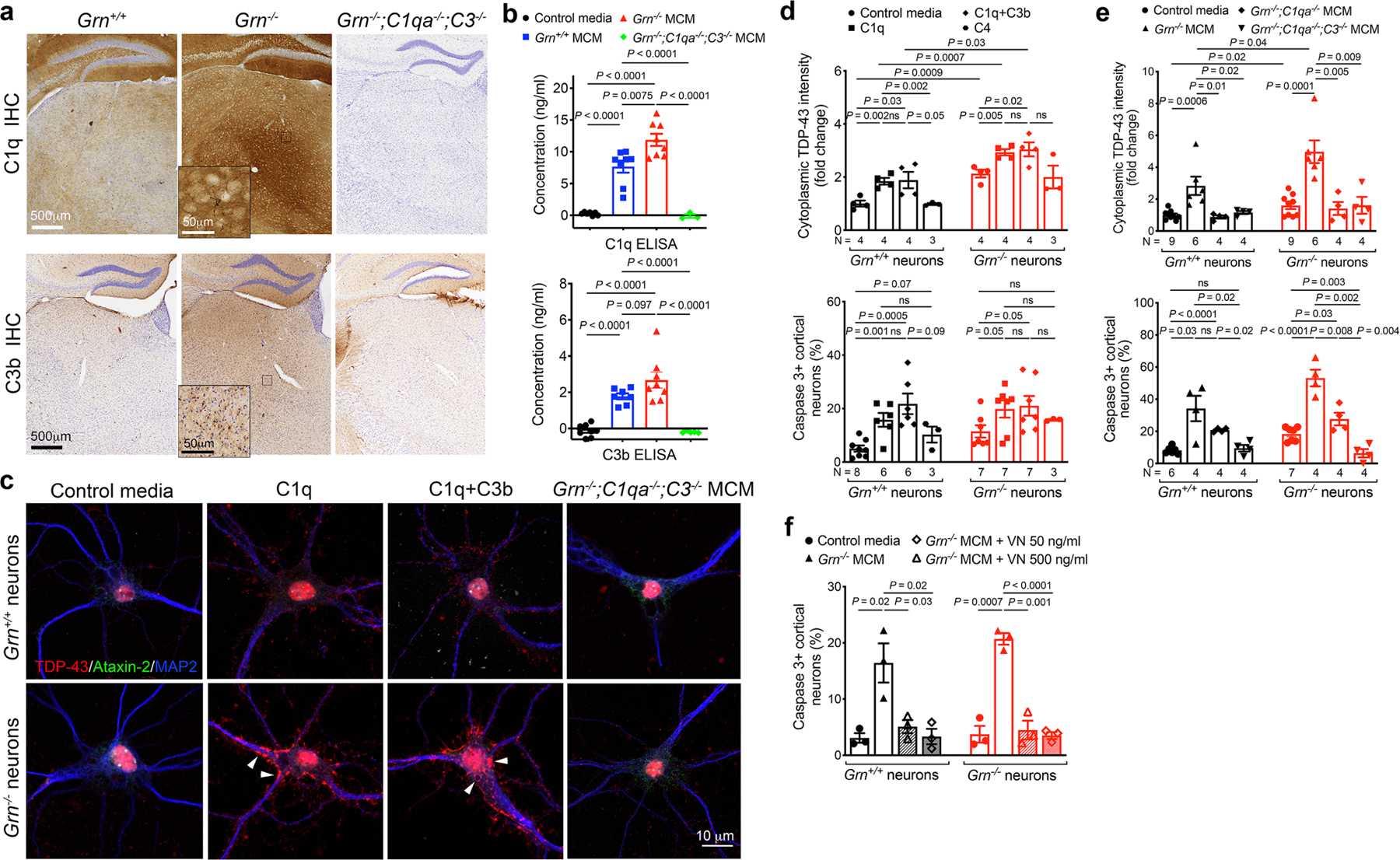
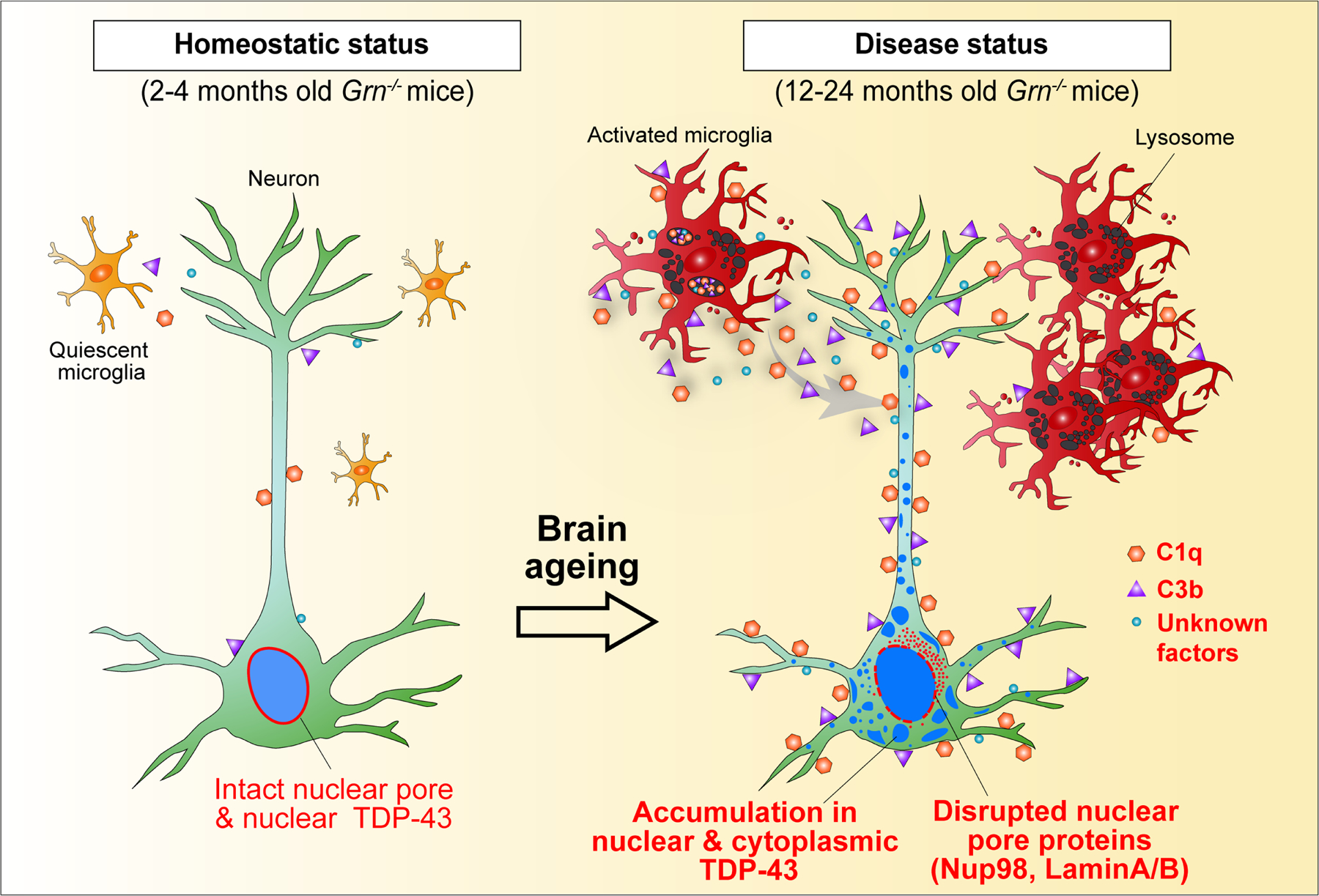
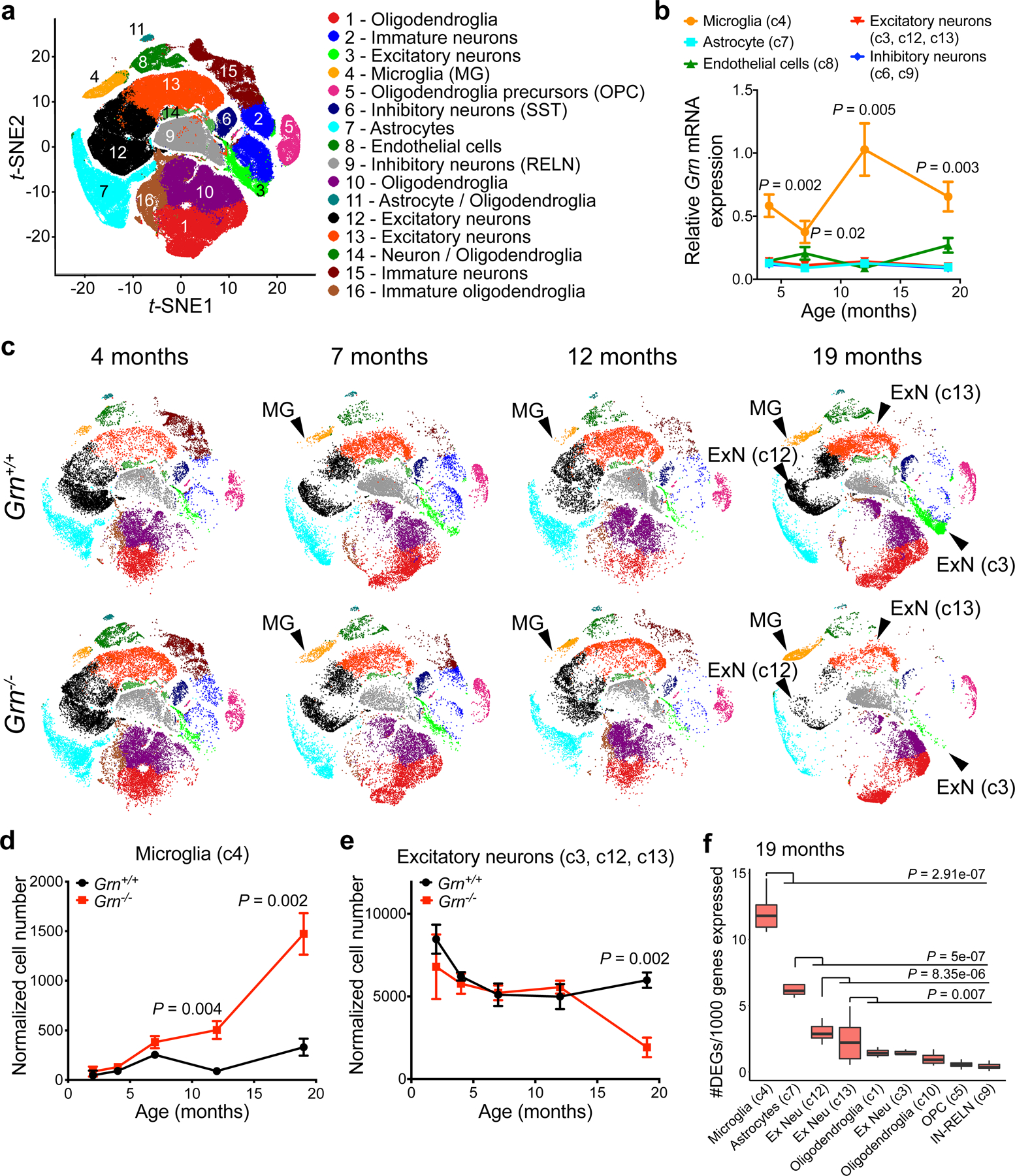
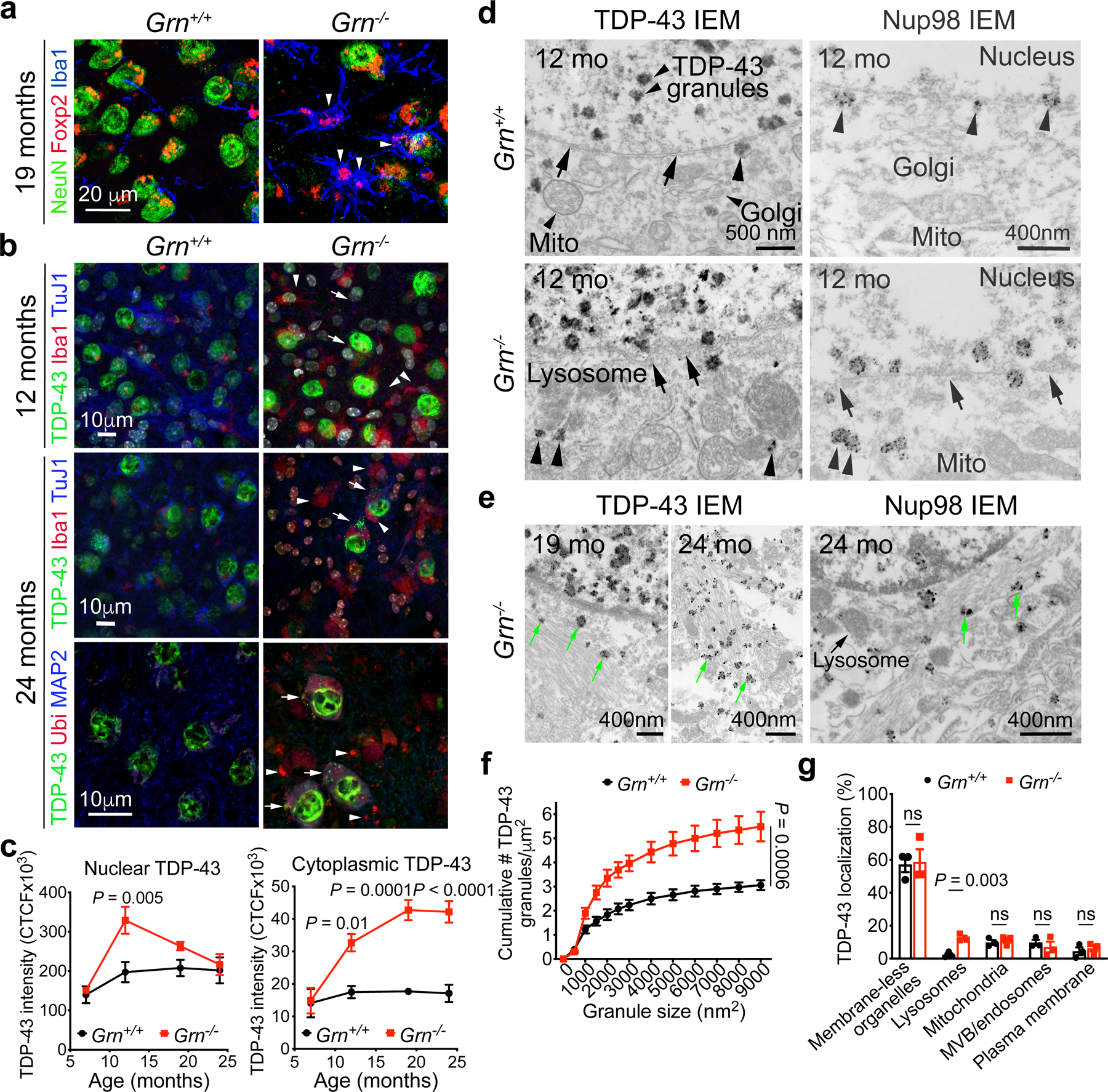
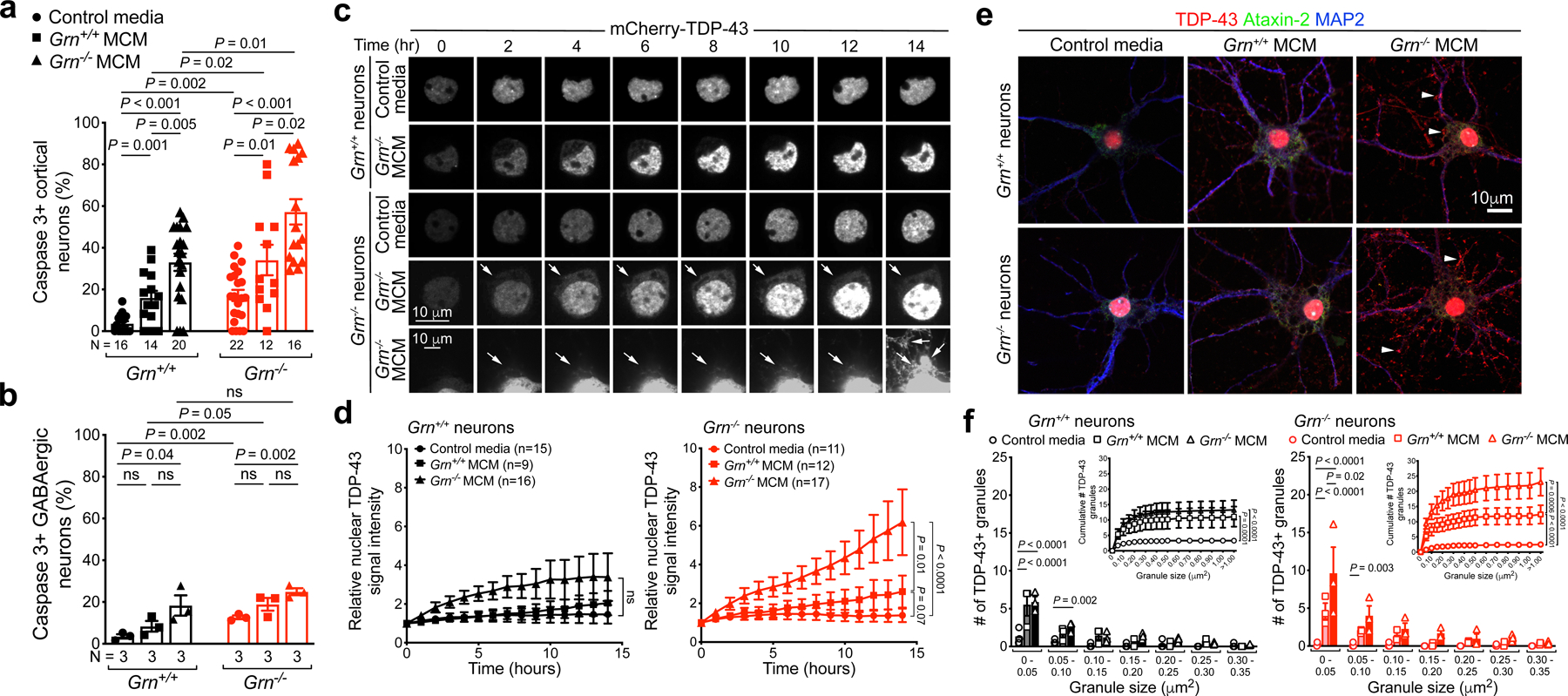
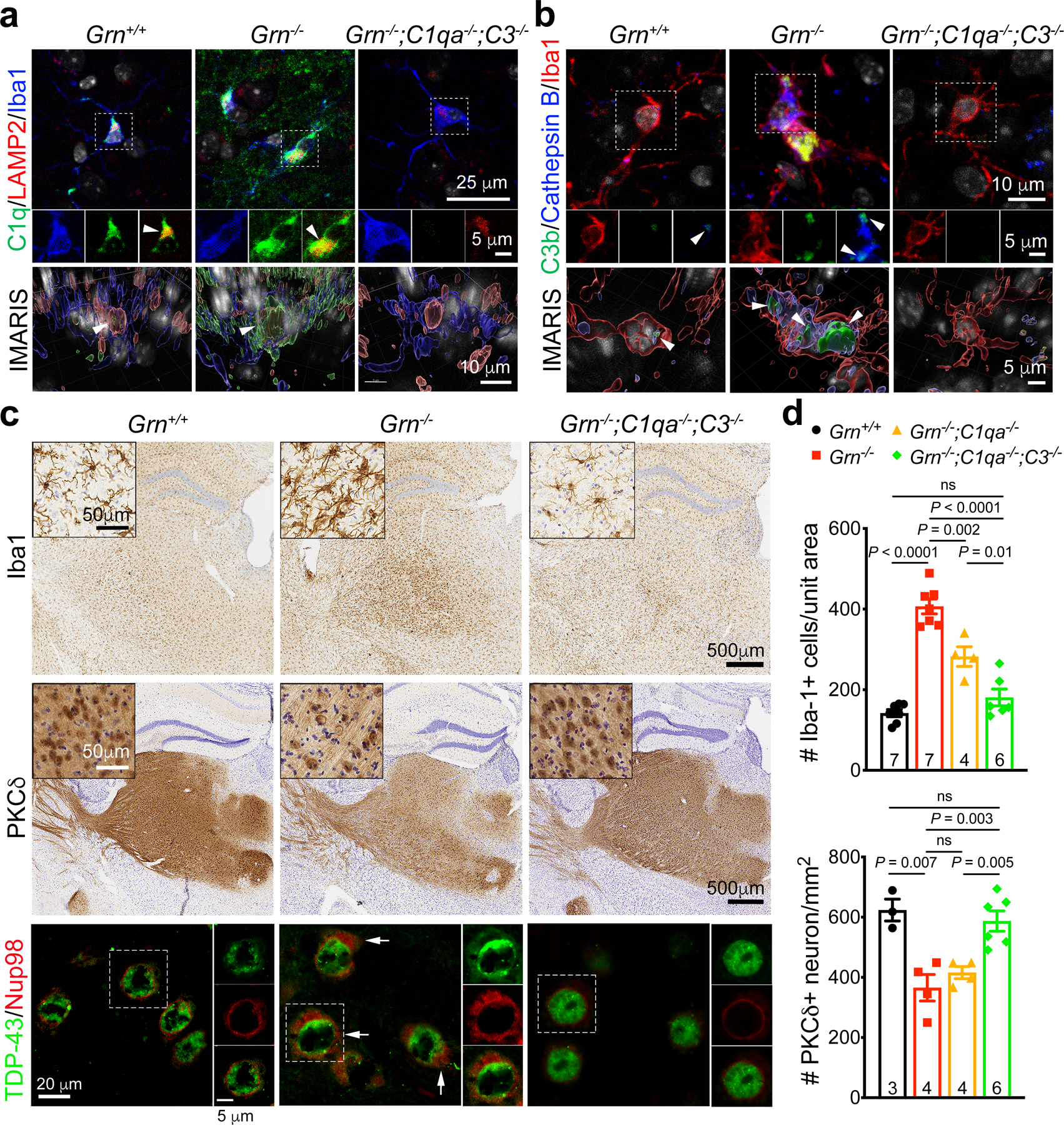
Aberrant aggregation of the RNA-binding protein TDP-43 in neurons is a hallmark of frontotemporal lobar degeneration caused by haploinsufficiency in the gene encoding progranulin1,2. However, the mechanism leading to TDP-43 proteinopathy remains unclear. Here we use single-nucleus RNA sequencing to show that progranulin deficiency promotes microglial transition from a homeostatic to a disease-specific state that causes endolysosomal dysfunction and neurodegeneration in mice. These defects persist even when Grn-/- microglia are cultured ex vivo. In addition, single-nucleus RNA sequencing reveals selective loss of excitatory neurons at disease end-stage, which is characterized by prominent nuclear and cytoplasmic TDP-43 granules and nuclear pore defects. Remarkably, conditioned media from Grn-/- microglia are sufficient to promote TDP-43 granule formation, nuclear pore defects and cell death in excitatory neurons via the complement activation pathway. Consistent with these results, deletion of the genes encoding C1qa and C3 mitigates microglial toxicity and rescues TDP-43 proteinopathy and neurodegeneration. These results uncover previously unappreciated contributions of chronic microglial toxicity to TDP-43 proteinopathy during neurodegeneration.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Additional References
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS107480/NS/NINDS NIH HHS/United States
- R35 NS097305/NS/NINDS NIH HHS/United States
- P30 AG062422/AG/NIA NIH HHS/United States
- R01 AG068290/AG/NIA NIH HHS/United States
- R01 CA231300/CA/NCI NIH HHS/United States
- K99 MH121534/MH/NIMH NIH HHS/United States
- R00 GM126136/GM/NIGMS NIH HHS/United States
- S10 RR026758/RR/NCRR NIH HHS/United States
- I01 BX002978/BX/BLRD VA/United States
- K99 GM126136/GM/NIGMS NIH HHS/United States
- U01 MH114825/MH/NIMH NIH HHS/United States
- U01 MH105989/MH/NIMH NIH HHS/United States
- R01 AG057462/AG/NIA NIH HHS/United States
- R01 GM124334/GM/NIGMS NIH HHS/United States
- R01 AA027074/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
