

Capping protein is dispensable for polarized actin network growth and actin-based motility
- PMID: 32868296
- PMCID: PMC7650241
- DOI: 10.1074/jbc.RA120.015009
Capping protein is dispensable for polarized actin network growth and actin-based motility
Abstract
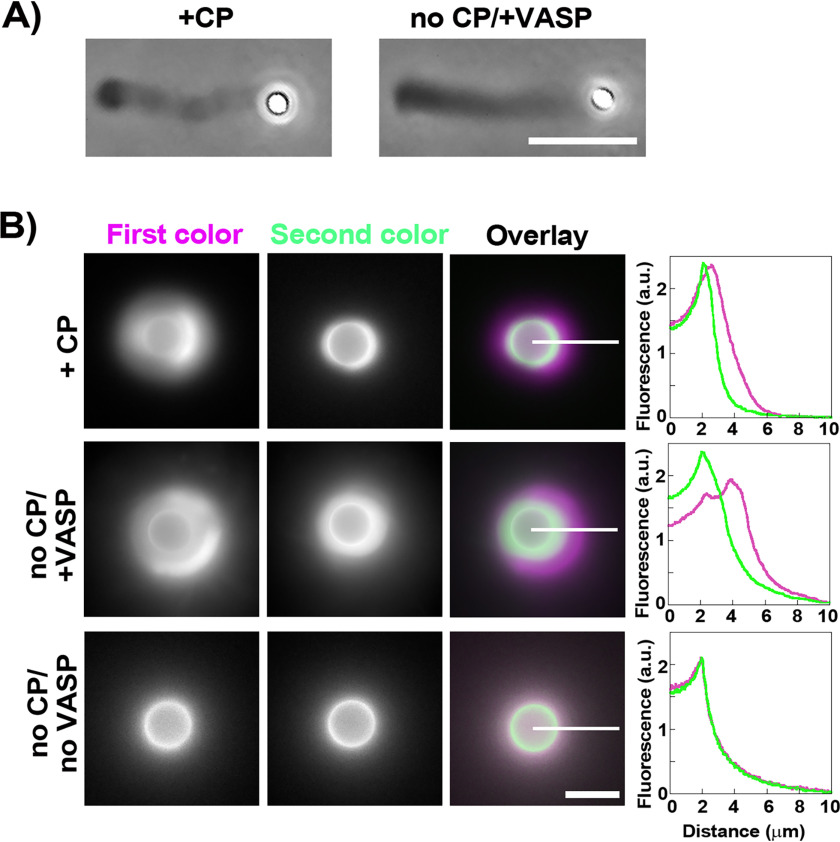
Heterodimeric capping protein (CP) binds the rapidly growing barbed ends of actin filaments and prevents the addition (or loss) of subunits. Capping activity is generally considered to be essential for actin-based motility induced by Arp2/3 complex nucleation. By stopping barbed end growth, CP favors nucleation of daughter filaments at the functionalized surface where the Arp2/3 complex is activated, thus creating polarized network growth, which is necessary for movement. However, here using an in vitro assay where Arp2/3 complex-based actin polymerization is induced on bead surfaces in the absence of CP, we produce robust polarized actin growth and motility. This is achieved either by adding the actin polymerase Ena/VASP or by boosting Arp2/3 complex activity at the surface. Another actin polymerase, the formin FMNL2, cannot substitute for CP, showing that polymerase activity alone is not enough to override the need for CP. Interfering with the polymerase activity of Ena/VASP, its surface recruitment or its bundling activity all reduce Ena/VASP's ability to maintain polarized network growth in the absence of CP. Taken together, our findings show that CP is dispensable for polarized actin growth and motility in situations where surface-directed polymerization is favored by whatever means over the growth of barbed ends in the network.
Keywords: Arp2/3 complex; Ena/VASP proteins; actin; actin polymerization; biophysics; capping protein; cell motility; cytoskeleton; in vitro systems.
© 2020 Abou-Ghali et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures


Similar articles
-
Capping protein regulatory cycle driven by CARMIL and V-1 may promote actin network assembly at protruding edges.Proc Natl Acad Sci U S A. 2014 May 13;111(19):E1970-9. doi: 10.1073/pnas.1313738111. Epub 2014 Apr 28. Proc Natl Acad Sci U S A. 2014. PMID: 24778263 Free PMC article.
-
A barbed end interference mechanism reveals how capping protein promotes nucleation in branched actin networks.Nat Commun. 2021 Sep 9;12(1):5329. doi: 10.1038/s41467-021-25682-5. Nat Commun. 2021. PMID: 34504078 Free PMC article.
-
Capping protein increases the rate of actin-based motility by promoting filament nucleation by the Arp2/3 complex.Cell. 2008 May 30;133(5):841-51. doi: 10.1016/j.cell.2008.04.011. Cell. 2008. PMID: 18510928 Free PMC article.
-
Capping protein regulators fine-tune actin assembly dynamics.Nat Rev Mol Cell Biol. 2014 Oct;15(10):677-89. doi: 10.1038/nrm3869. Epub 2014 Sep 10. Nat Rev Mol Cell Biol. 2014. PMID: 25207437 Free PMC article. Review.
-
Ena/VASP proteins: regulators of the actin cytoskeleton and cell migration.Annu Rev Cell Dev Biol. 2003;19:541-64. doi: 10.1146/annurev.cellbio.19.050103.103356. Annu Rev Cell Dev Biol. 2003. PMID: 14570581 Review.
Cited by
-
Prediction of the essential intermolecular contacts for side-binding of VASP on F-actin.Cytoskeleton (Hoboken). 2024 Aug;81(8):382-392. doi: 10.1002/cm.21864. Epub 2024 Apr 22. Cytoskeleton (Hoboken). 2024. PMID: 38647032 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
