

Sterile Spikelets Contribute to Yield in Sorghum and Related Grasses
- PMID: 32873633
- PMCID: PMC7610286
- DOI: 10.1105/tpc.20.00424
Sterile Spikelets Contribute to Yield in Sorghum and Related Grasses
Abstract
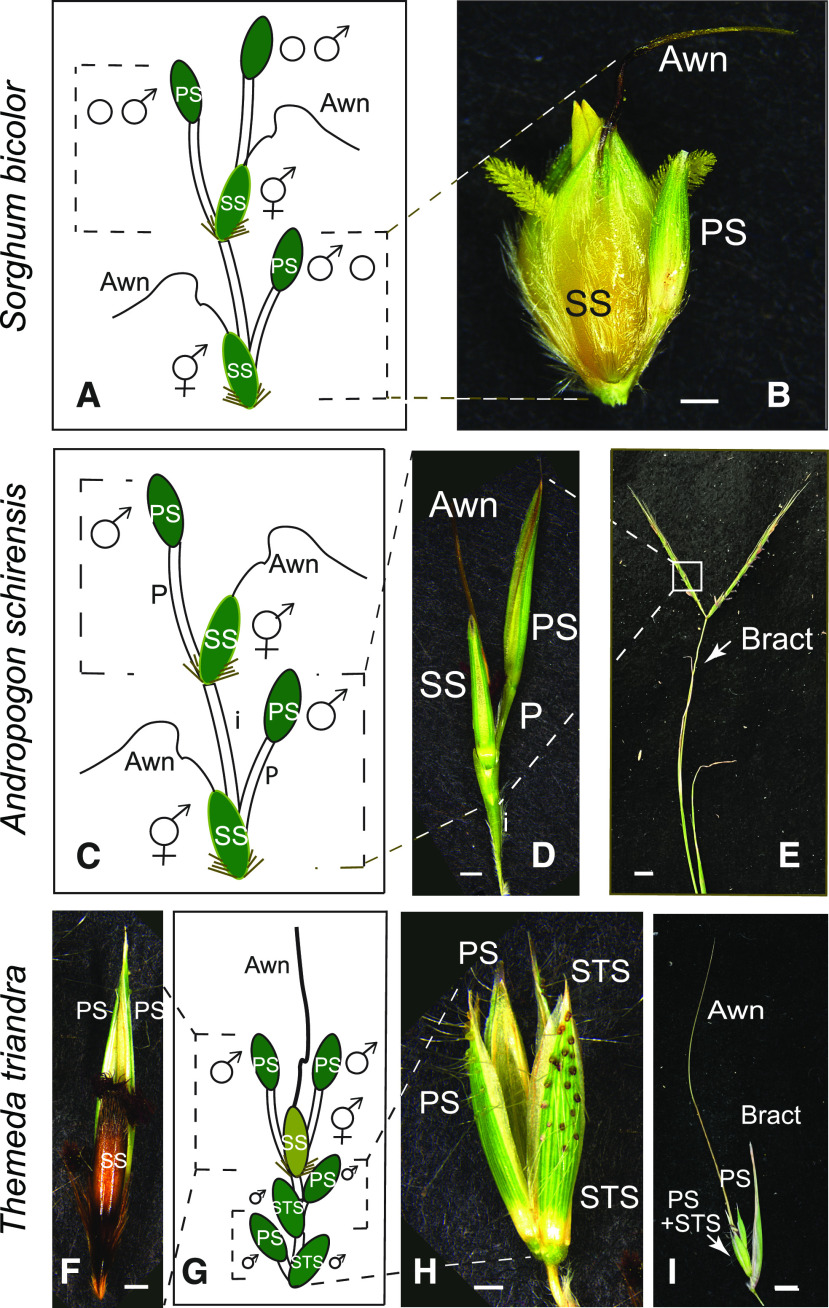
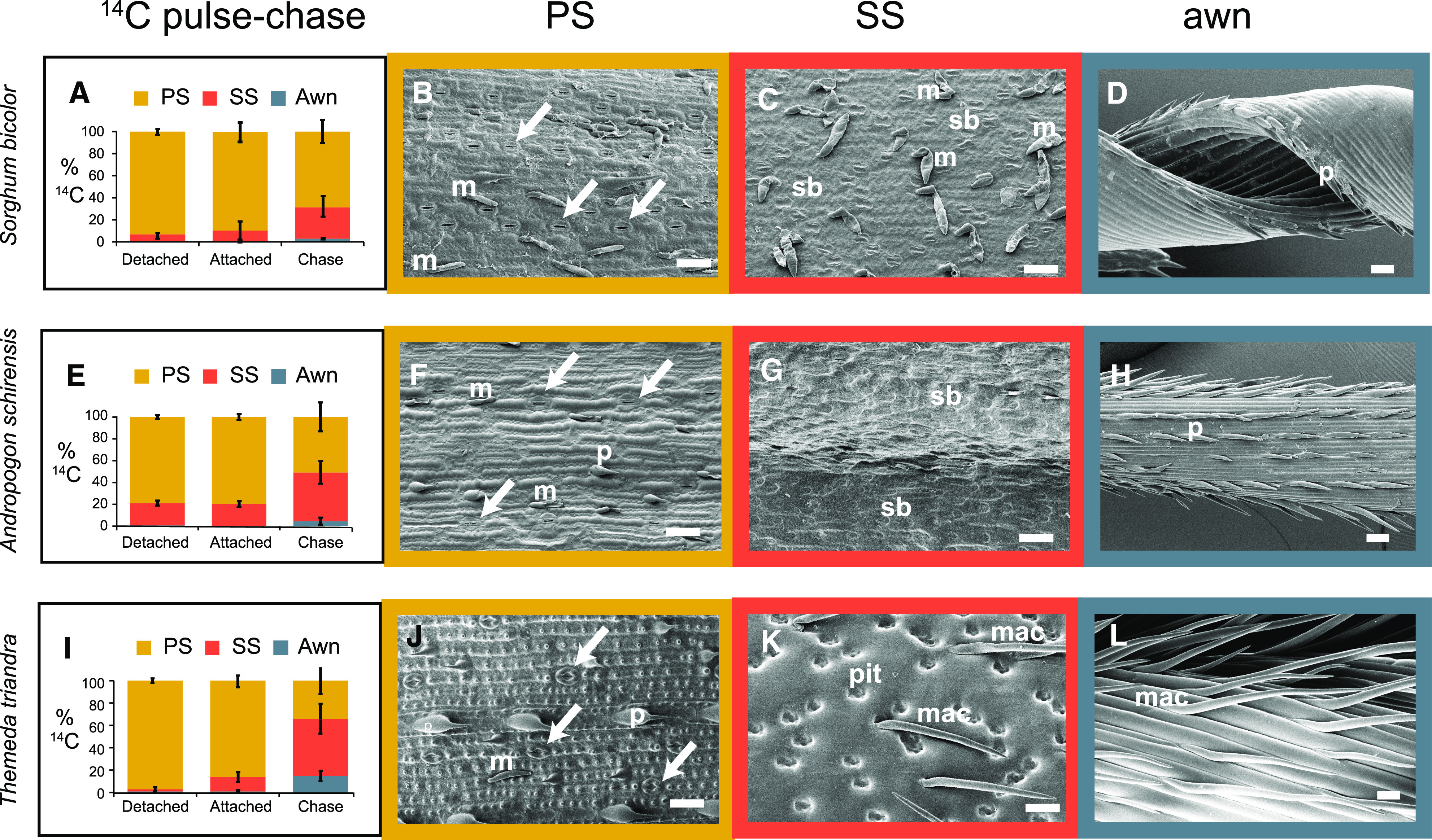
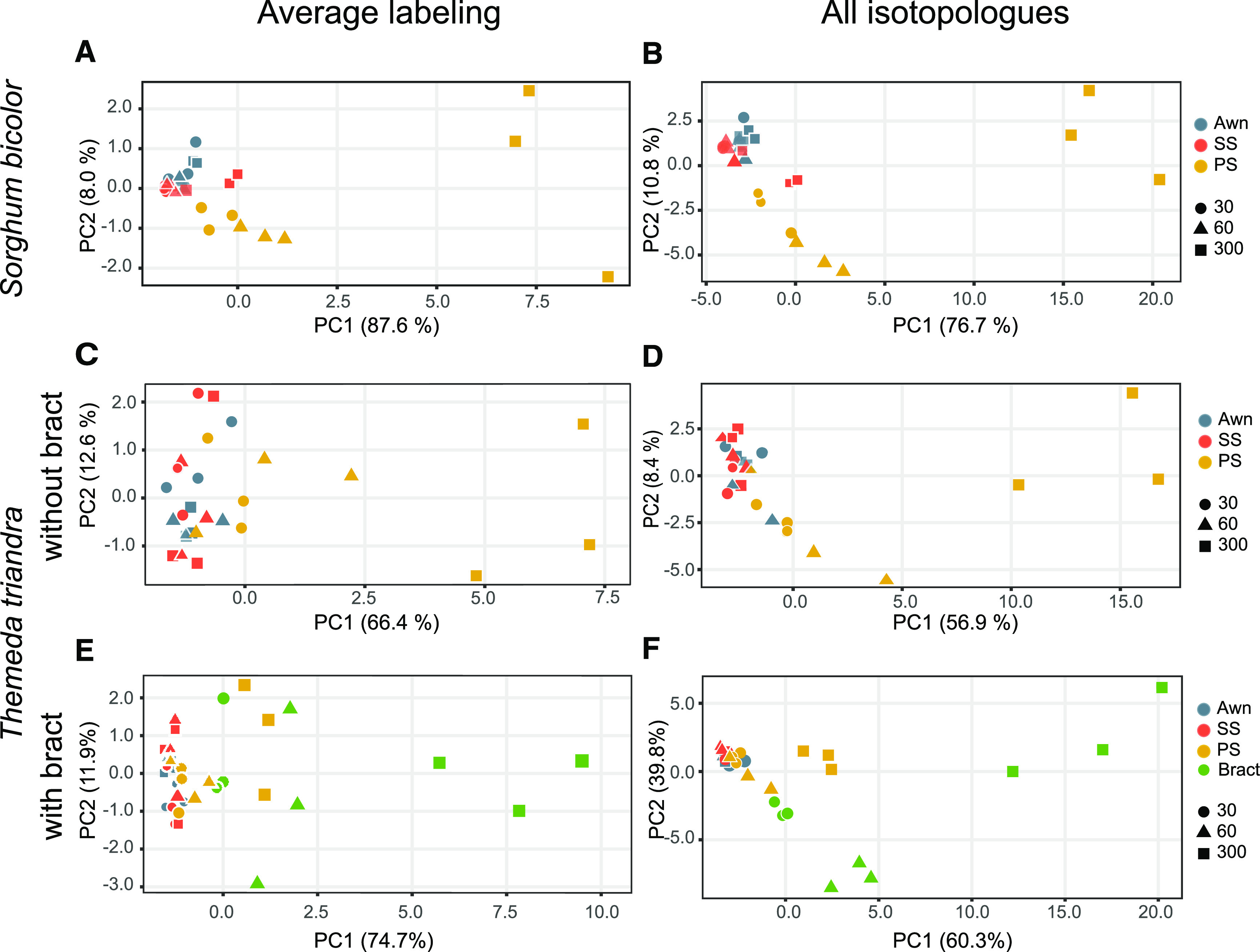
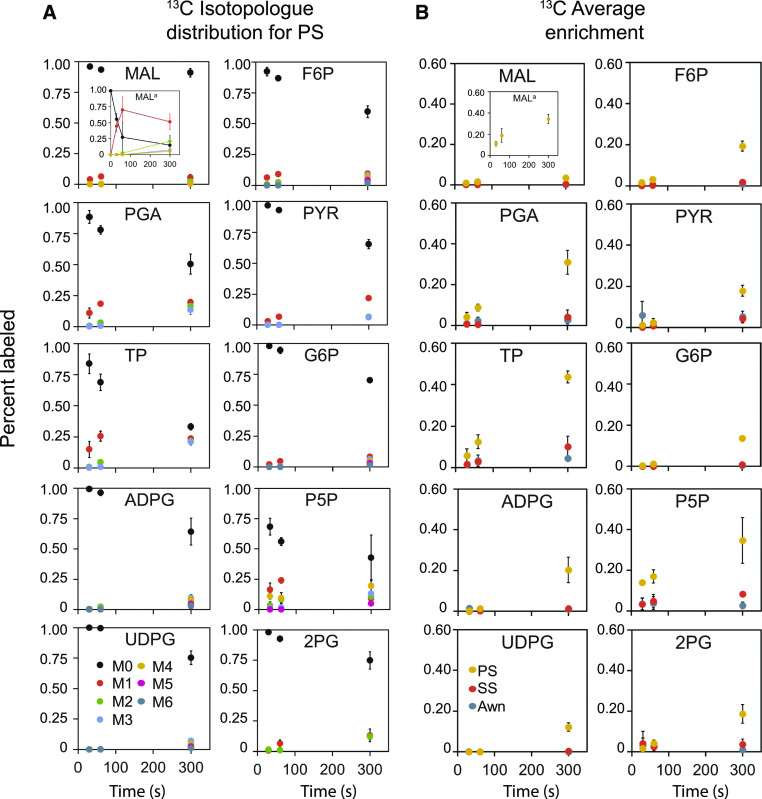
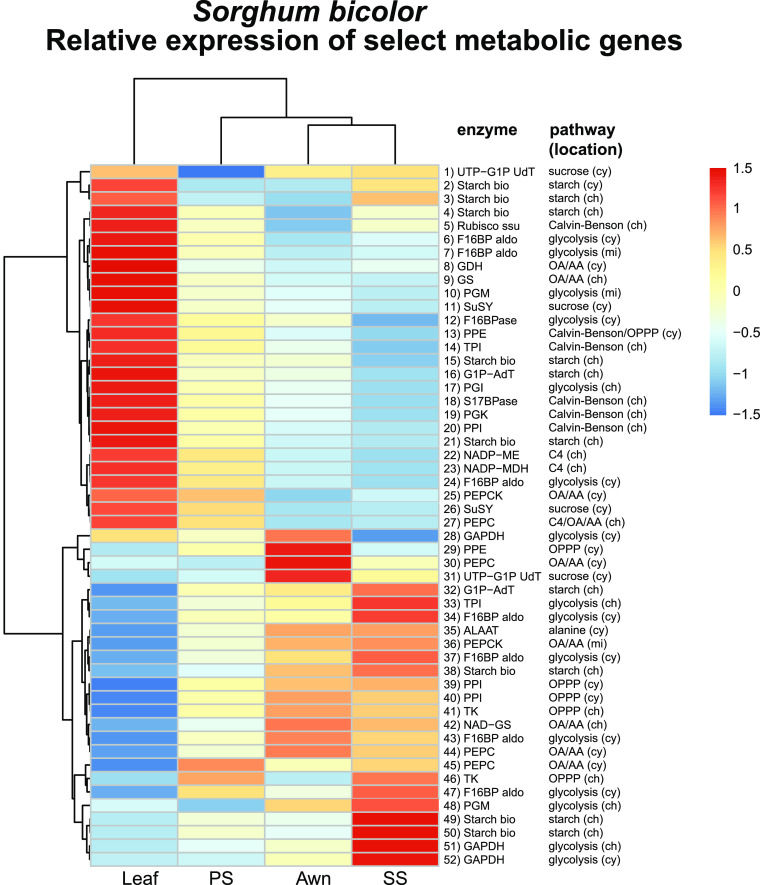
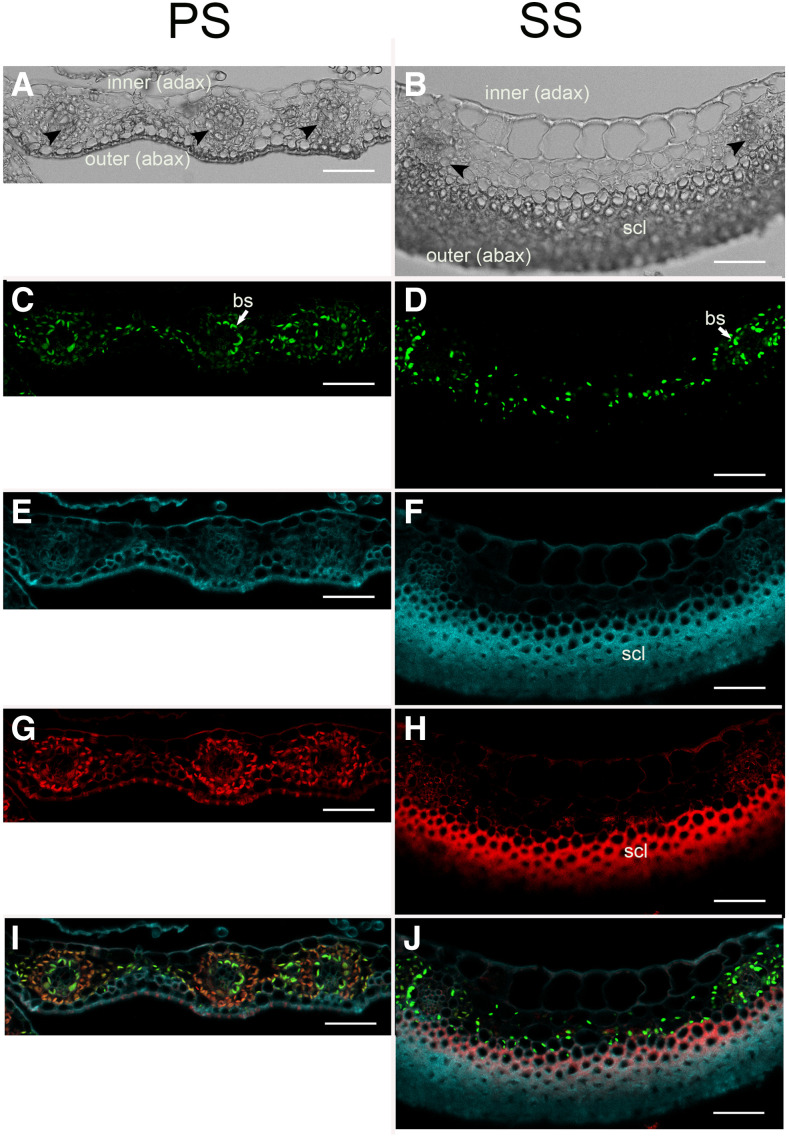
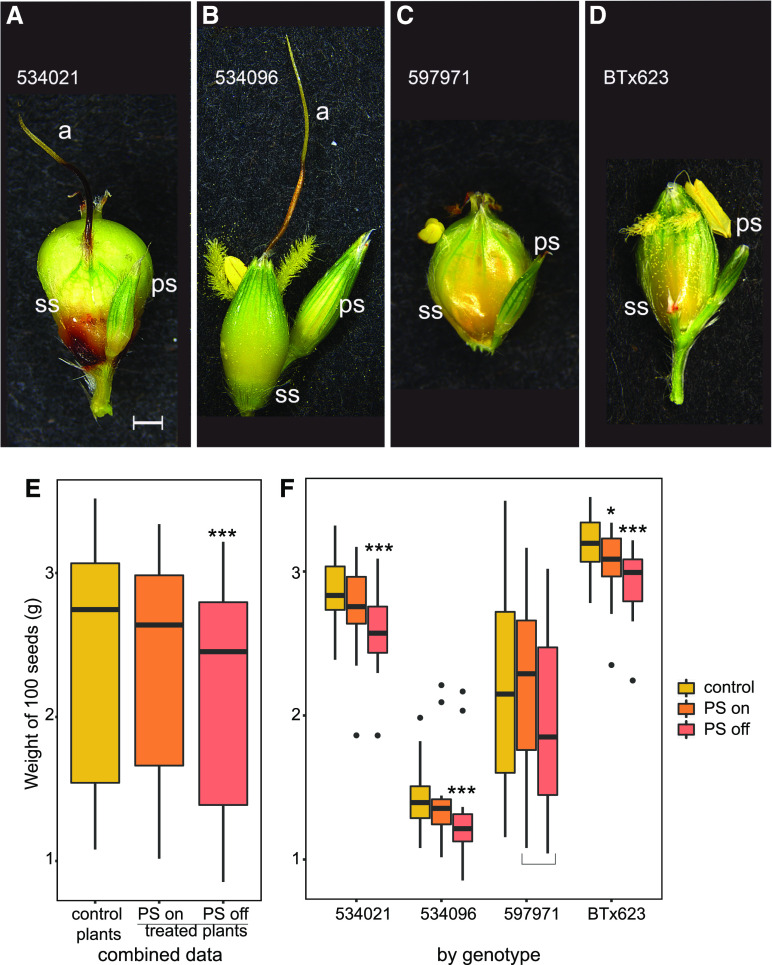
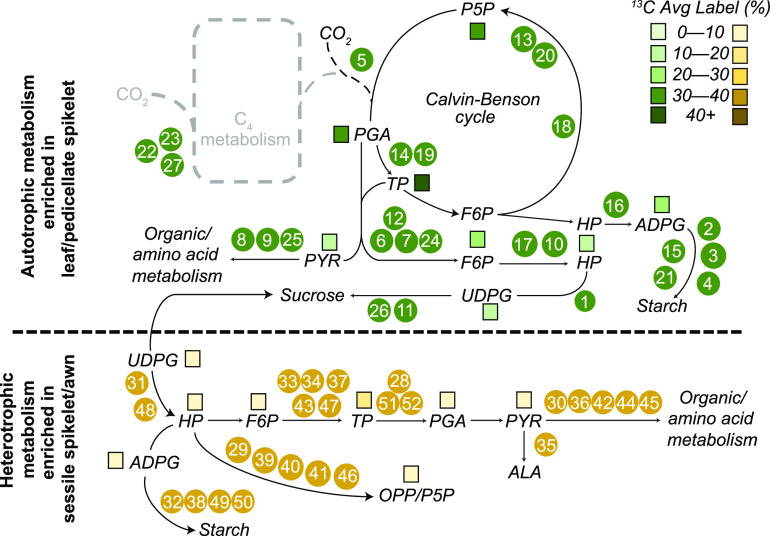
Sorghum (Sorghum bicolor) and its relatives in the grass tribe Andropogoneae bear their flowers in pairs of spikelets in which one spikelet (seed-bearing or sessile spikelet [SS]) of the pair produces a seed and the other is sterile or male (staminate). This division of function does not occur in other major cereals such as wheat (Triticum aestivum) or rice (Oryza sativa). Additionally, one bract of the SS spikelet often produces a long extension, the awn, that is in the same position as, but independently derived from, that of wheat and rice. The function of the sterile spikelet is unknown and that of the awn has not been tested in Andropogoneae. We used radioactive and stable isotopes of carbon, RNA sequencing of metabolically important enzymes, and immunolocalization of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) to show that the sterile spikelet assimilates carbon, which is translocated to the largely heterotrophic SS. The awn shows no evidence of photosynthesis. These results apply to distantly related species of Andropogoneae. Removal of sterile spikelets in sorghum significantly decreases seed weight (yield) by ∼9%. Thus, the sterile spikelet, but not the awn, affects yield in the cultivated species and fitness in the wild species.
© 2020 American Society of Plant Biologists. All rights reserved.
Figures


Comment in
-
Gains in Grain Yield: A Pair of Spikelets Makes All the Difference, Even When One Is Sterile.Plant Cell. 2020 Nov;32(11):3378-3379. doi: 10.1105/tpc.20.00667. Epub 2020 Sep 3. Plant Cell. 2020. PMID: 32883712 Free PMC article. No abstract available.
References
-
- Abramoff M.D., Magalhaes P.J., Ram S.J.(2004). Image processing with ImageJ. Biophoton. Int. 11: 36–42.
-
- Allen D.K., Ohlrogge J.B., Shachar-Hill Y.(2009). The role of light in soybean seed filling metabolism. Plant J. 58: 220–234. - PubMed
-
- Antlfinger A., Wendel L.(1997). Reproductive effort and floral photosynthesis in Spiranthes cernua (Orchidaceae). Am. J. Bot. 84: 769–780. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
