

Use of CRISPR-Cas9 To Target Homologous Recombination Limits Transformation-Induced Genomic Changes in Candida albicans
- PMID: 32878930
- PMCID: PMC7471004
- DOI: 10.1128/mSphere.00620-20
Use of CRISPR-Cas9 To Target Homologous Recombination Limits Transformation-Induced Genomic Changes in Candida albicans
Abstract
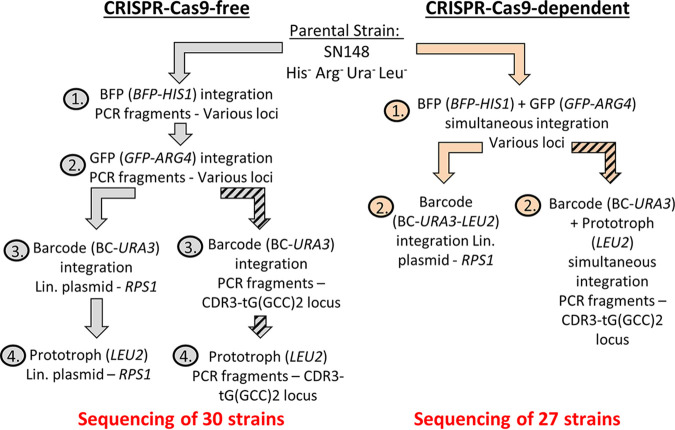
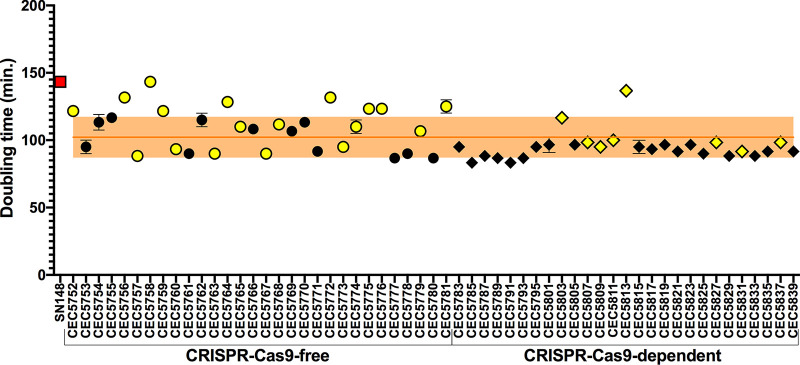
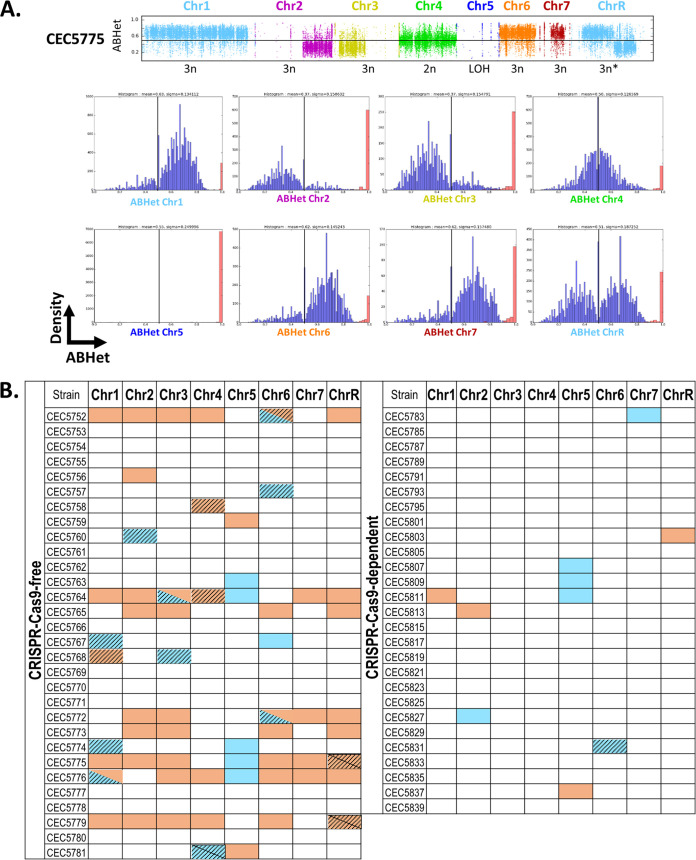
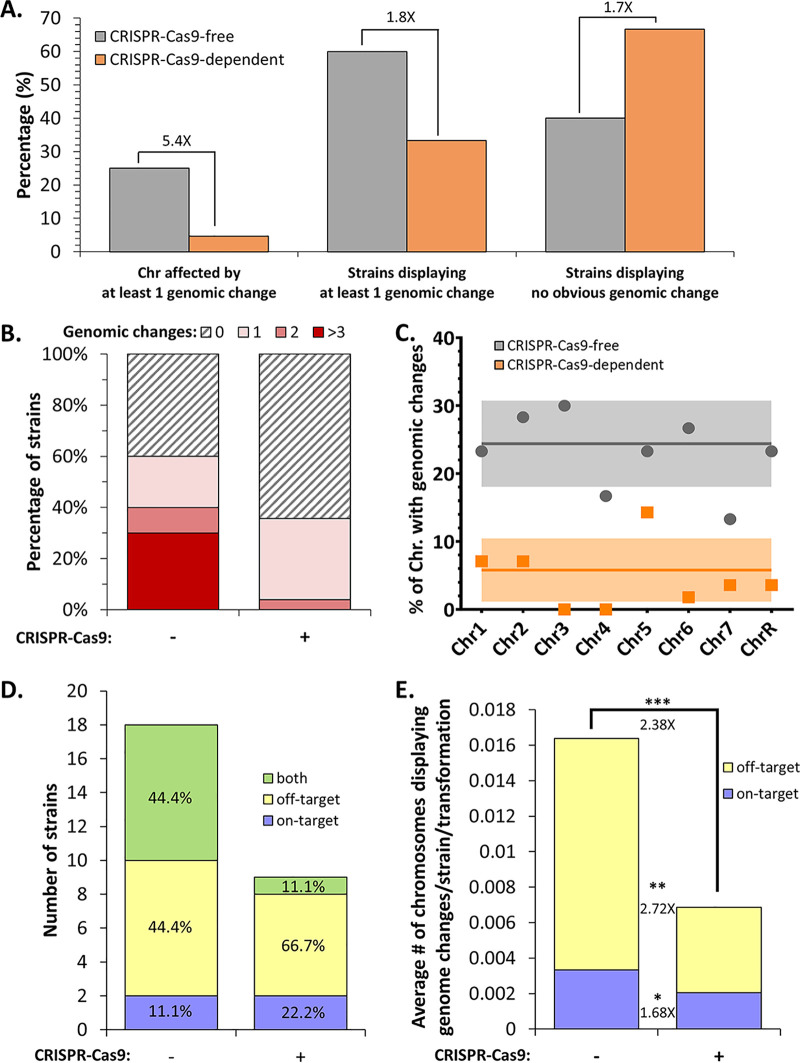
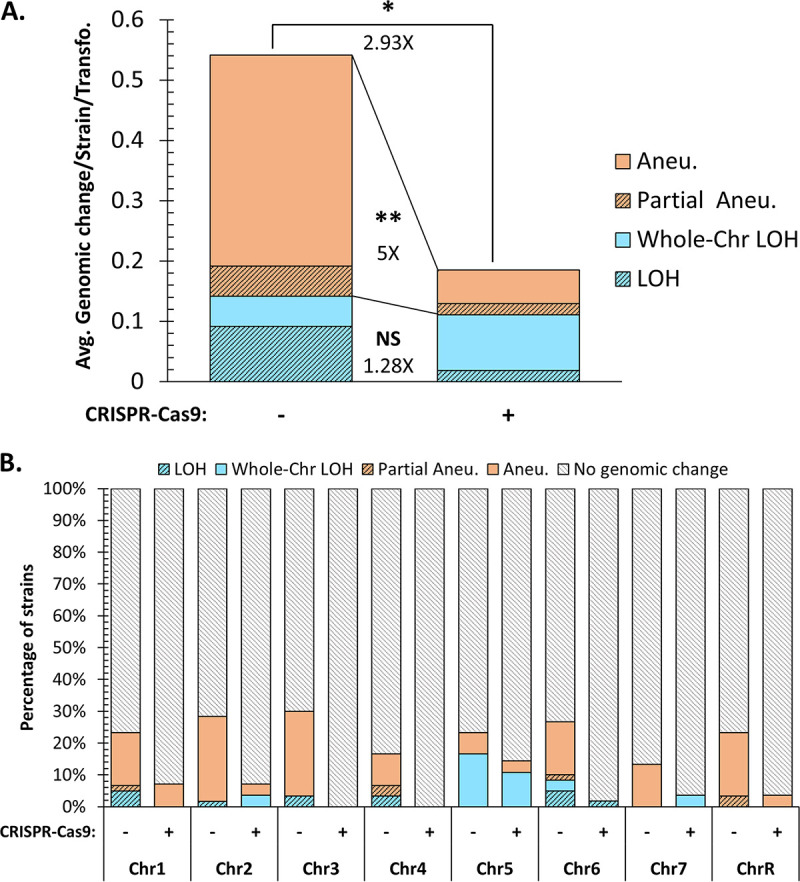
Most of our knowledge relating to molecular mechanisms of human fungal pathogenesis in Candida albicans relies on reverse genetics approaches, requiring strain engineering. DNA-mediated transformation of C. albicans has been described as highly mutagenic, potentially accentuated by the organism's genome plasticity, including the acquisition of genomic rearrangements, notably upon exposure to stress. The advent of CRISPR-Cas9 has vastly accelerated the process of genetically modifying strains, especially in diploid (such as C. albicans) and polyploid organisms. The effects of unleashing this nuclease within the genome of C. albicans are unknown, although several studies in other organisms report Cas9-associated toxicity and off-target DNA breaks. Upon the construction of a C. albicans strain collection, we took the opportunity to compare strains which were constructed using CRISPR-Cas9-free and CRISPR-Cas9-dependent transformation strategies, by quantifying and describing transformation-induced loss-of-heterozygosity and hyperploidy events. Our analysis of 57 strains highlights the mutagenic effects of transformation in C. albicans, regardless of the transformation protocol, but also underscores interesting differences in terms of genomic changes between strains obtained using different transformation protocols. Indeed, although strains constructed using the CRISPR-Cas9-free transformation method display numerous concomitant genomic changes randomly distributed throughout their genomes, the use of CRISPR-Cas9 leads to a reduced overall number of genome changes, particularly hyperploidies. Overall, in addition to facilitating strain construction by reducing the number of transformation steps, the CRISPR-Cas9-dependent transformation strategy in C. albicans appears to limit transformation-associated genome changes.IMPORTANCE Genome editing is essential to nearly all research studies aimed at gaining insight into the molecular mechanisms underlying various biological processes, including those in the opportunistic pathogen Candida albicans The adaptation of the CRISPR-Cas9 system greatly facilitates genome engineering in many organisms. However, our understanding of the effects of CRISPR-Cas9 technology on the biology of C. albicans is limited. In this study, we sought to compare the extents of transformation-induced genomic changes within strains engineered using CRISPR-Cas9-free and CRISPR-Cas9-dependent transformation methods. CRISPR-Cas9-dependent transformation allows one to simultaneously target both homologs and, importantly, appears less mutagenic in C. albicans, since strains engineered using CRISPR-Cas9 display an overall decrease in concomitant genomic changes.
Keywords: CRISPR-Cas9; Candida albicans; genome rearrangements.
Copyright © 2020 Marton et al.
Figures
References
-
- Legrand M, Bachellier-Bassi S, Lee KK, Chaudhari Y, Tournu H, Arbogast L, Boyer H, Chauvel M, Cabral V, Maufrais C, Nesseir A, Maslanka I, Permal E, Rossignol T, Walker LA, Zeidler U, Znaidi S, Schoeters F, Majgier C, Julien RA, Ma L, Tichit M, Bouchier C, Dijck PV, Munro CA, D’Enfert C. 2018. Generating genomic platforms to study Candida albicans pathogenesis. Nucleic Acids Res 46:6935–6949. doi:10.1093/nar/gky594. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
