

Mutant thermal proteome profiling for characterization of missense protein variants and their associated phenotypes within the proteome
- PMID: 32878984
- PMCID: PMC7705321
- DOI: 10.1074/jbc.RA120.014576
Mutant thermal proteome profiling for characterization of missense protein variants and their associated phenotypes within the proteome
Abstract
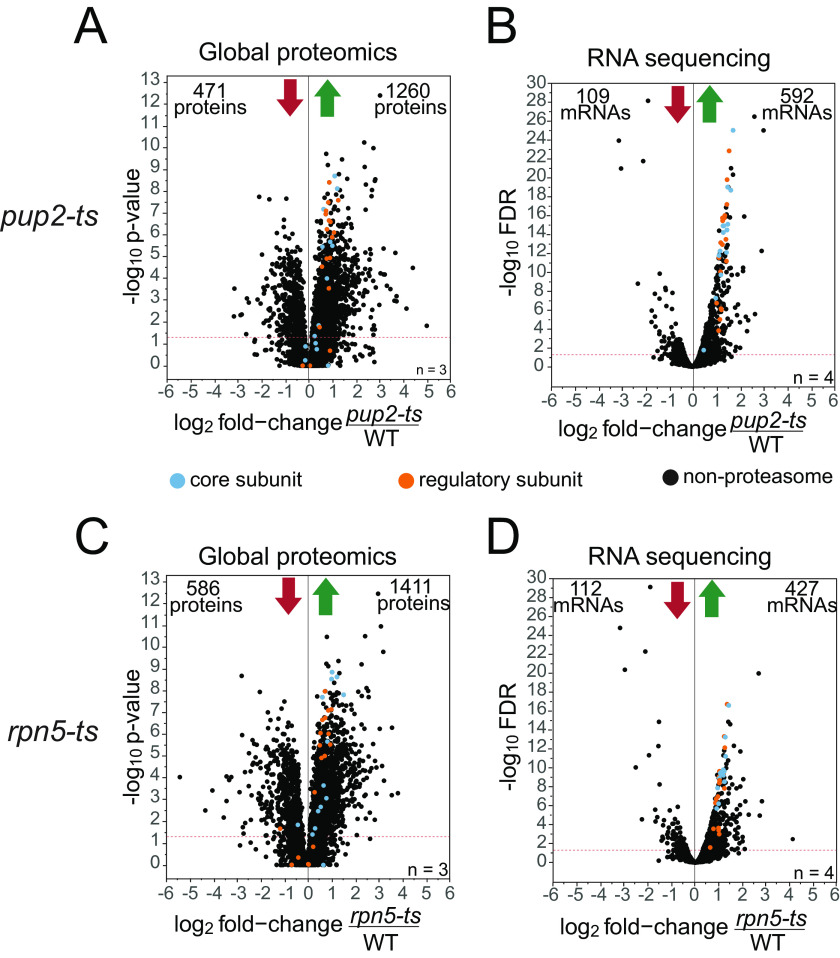
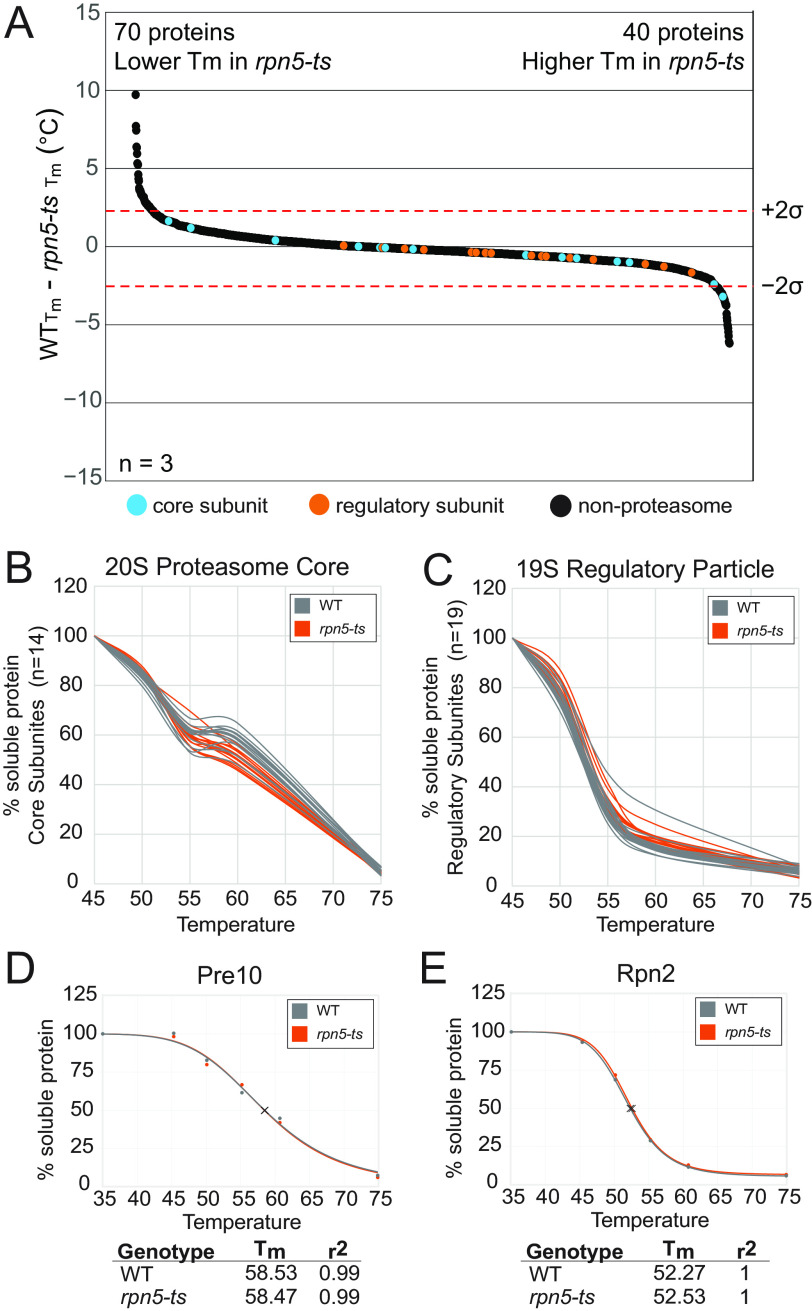
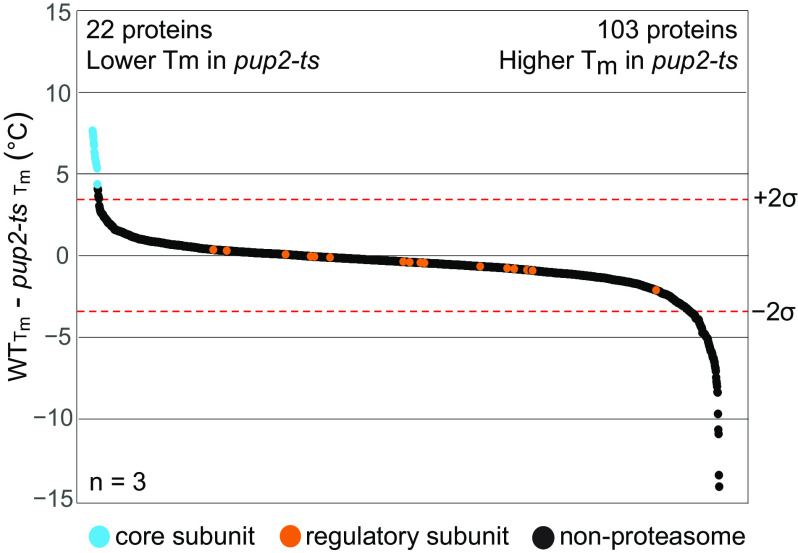
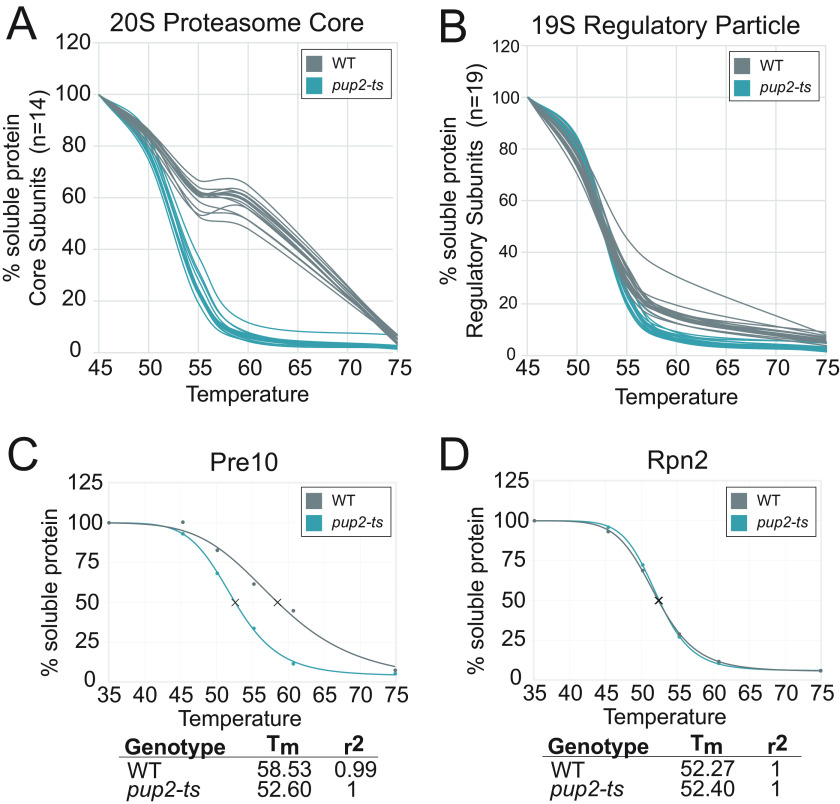
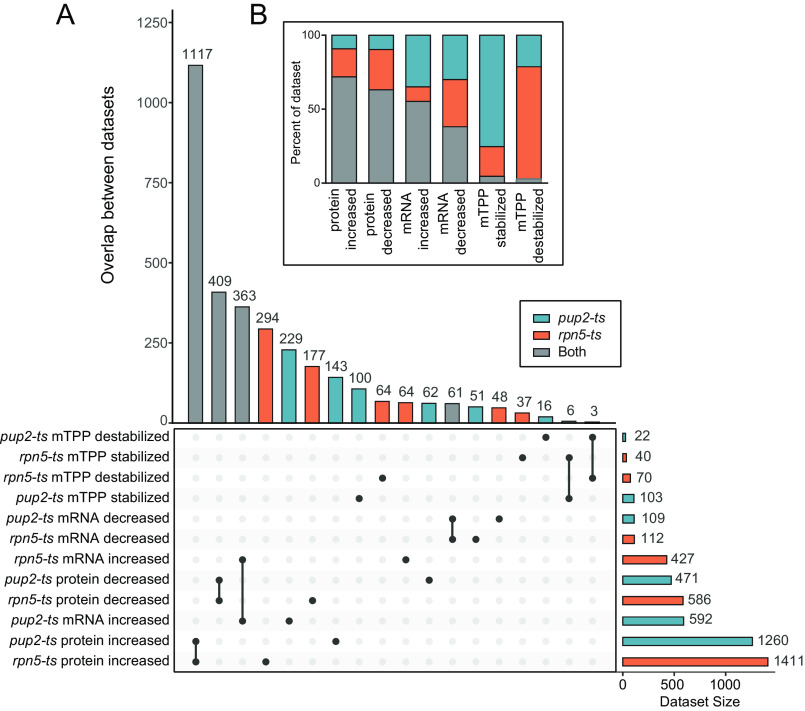
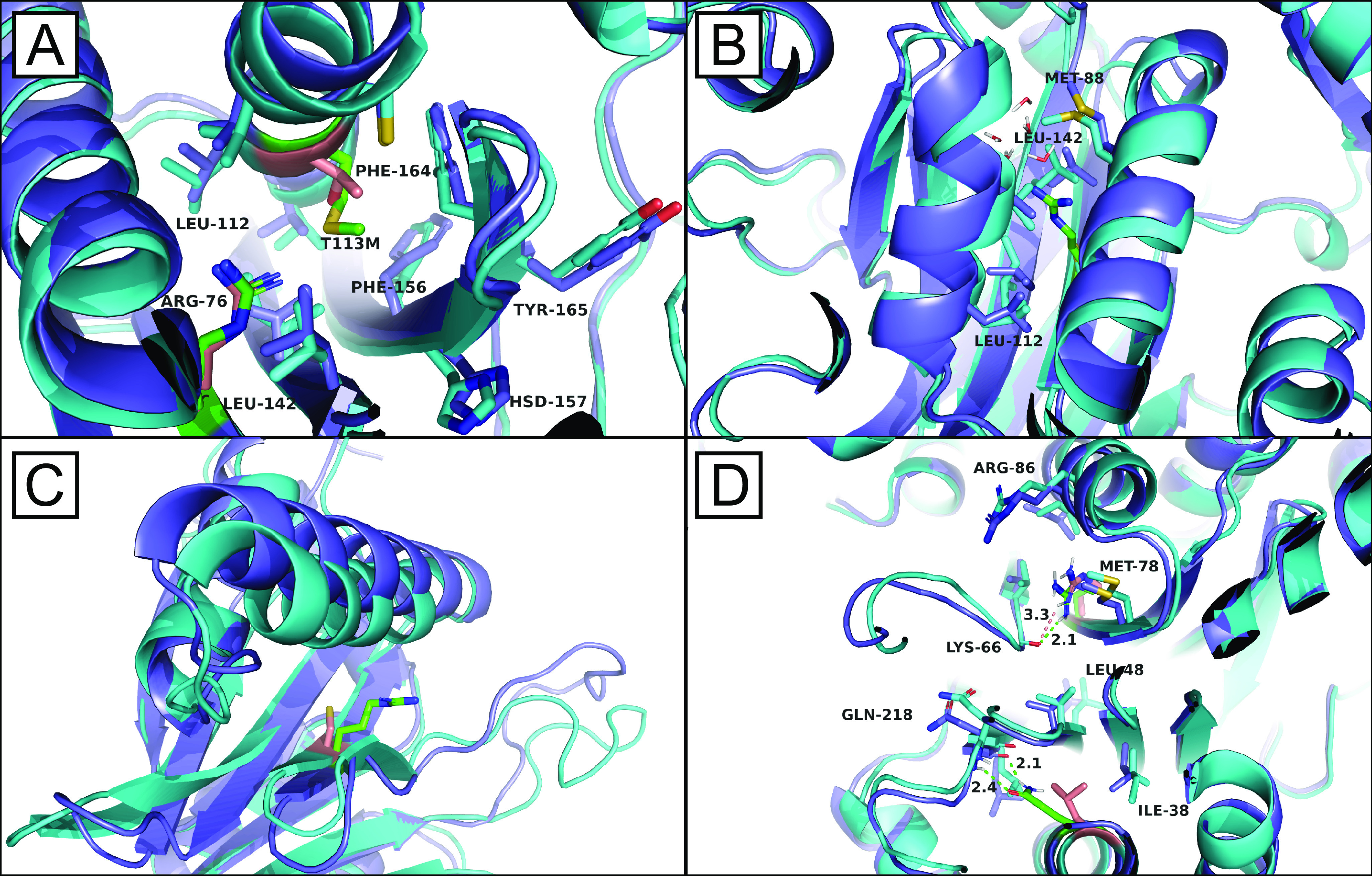
Temperature-sensitive (TS) missense mutants have been foundational for characterization of essential gene function. However, an unbiased approach for analysis of biochemical and biophysical changes in TS missense mutants within the context of their functional proteomes is lacking. We applied MS-based thermal proteome profiling (TPP) to investigate the proteome-wide effects of missense mutations in an application that we refer to as
Keywords: mass spectrometry; missense variant; mutant; proteasome; protein complex; protein stability; protein structure; protein-protein interaction; proteomics; systems biology; temperature-sensitive; thermal profiling.
© 2020 Peck Justice et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures


References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
