

Plasmacytoid dendritic cells cross-prime naive CD8 T cells by transferring antigen to conventional dendritic cells through exosomes
- PMID: 32879009
- PMCID: PMC7519282
- DOI: 10.1073/pnas.2002345117
Plasmacytoid dendritic cells cross-prime naive CD8 T cells by transferring antigen to conventional dendritic cells through exosomes
Abstract
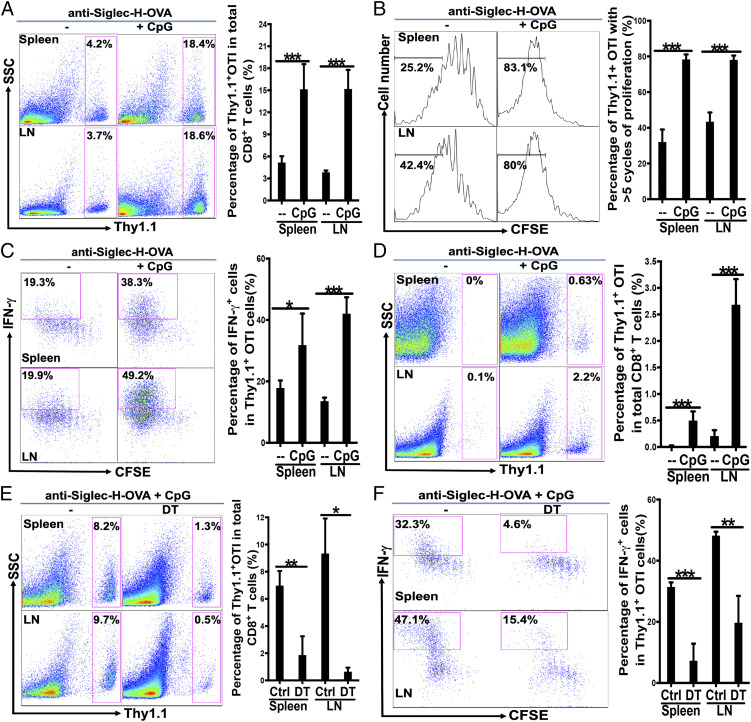
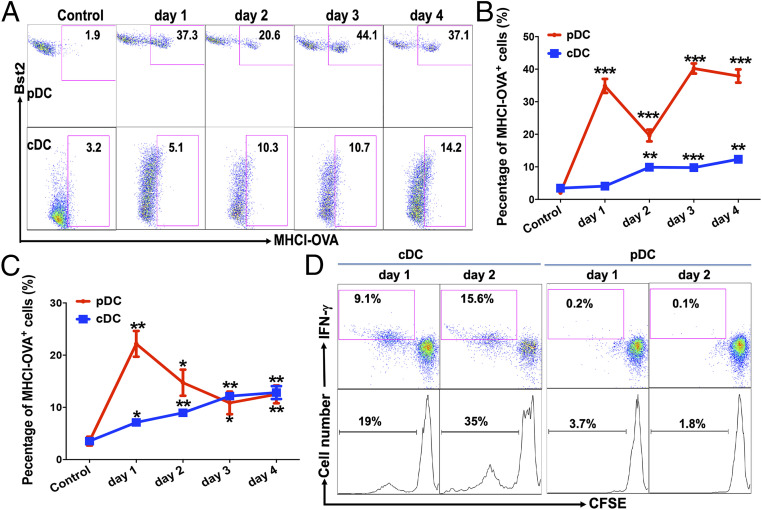
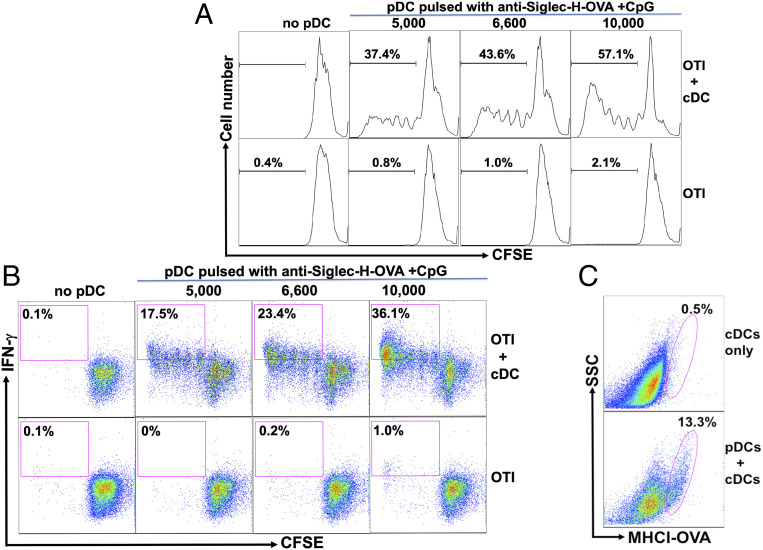
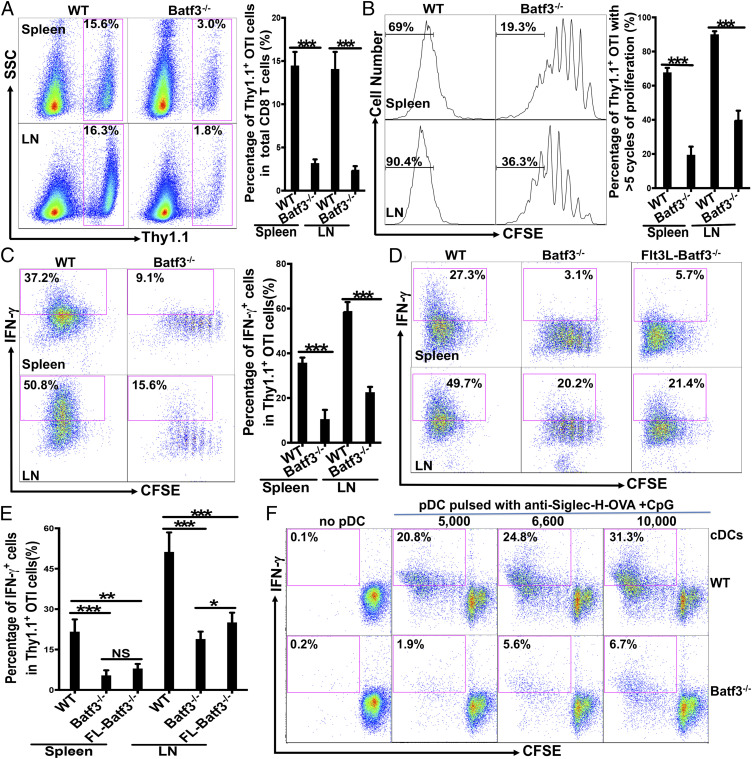
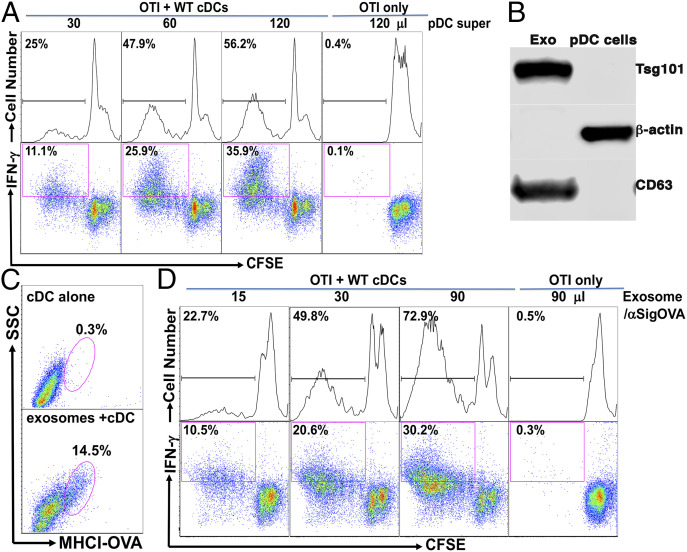
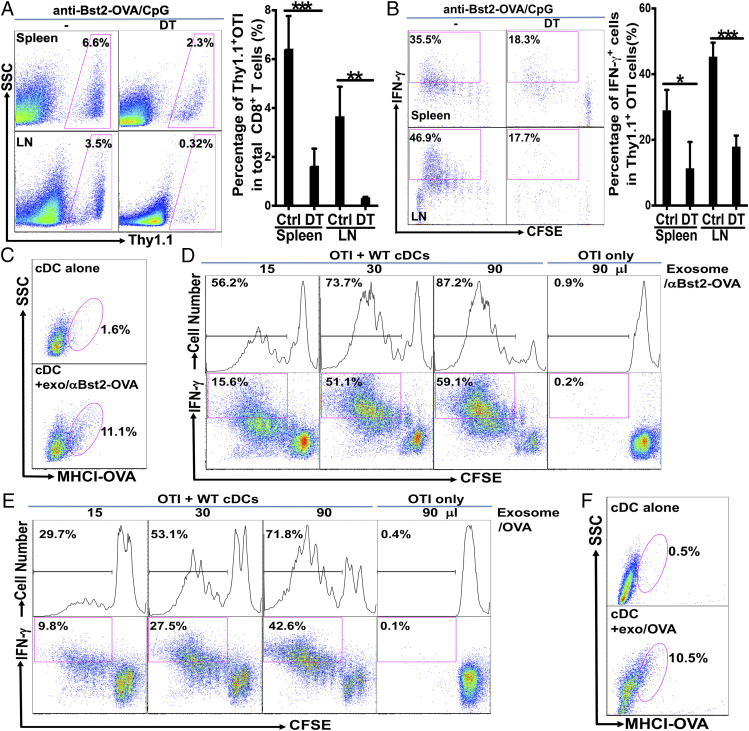
Although plasmacytoid dendritic cells (pDCs) have been shown to play a critical role in generating viral immunity and promoting tolerance to suppress antitumor immunity, whether and how pDCs cross-prime CD8 T cells in vivo remain controversial. Using a pDC-targeted vaccine model to deliver antigens specifically to pDCs, we have demonstrated that pDC-targeted vaccination led to strong cross-priming and durable CD8 T cell immunity. Surprisingly, cross-presenting pDCs required conventional DCs (cDCs) to achieve cross-priming in vivo by transferring antigens to cDCs. Taking advantage of an in vitro system where only pDCs had access to antigens, we further demonstrated that cross-presenting pDCs were unable to efficiently prime CD8 T cells by themselves, but conferred antigen-naive cDCs the capability of cross-priming CD8 T cells by transferring antigens to cDCs. Although both cDC1s and cDC2s exhibited similar efficiency in acquiring antigens from pDCs, cDC1s but not cDC2s were required for cross-priming upon pDC-targeted vaccination, suggesting that cDC1s played a critical role in pDC-mediated cross-priming independent of their function in antigen presentation. Antigen transfer from pDCs to cDCs was mediated by previously unreported pDC-derived exosomes (pDCexos), that were also produced by pDCs under various conditions. Importantly, all these pDCexos primed naive antigen-specific CD8 T cells only in the presence of bystander cDCs, similarly to cross-presenting pDCs, thus identifying pDCexo-mediated antigen transfer to cDCs as a mechanism for pDCs to achieve cross-priming. In summary, our data suggest that pDCs employ a unique mechanism of pDCexo-mediated antigen transfer to cDCs for cross-priming.
Keywords: antigen transfer; conventional dendritic cells; cross-priming; exosomes; plasmacytoid dendritic cells.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
DC-Based Vaccines for Cancer Immunotherapy.Vaccines (Basel). 2020 Nov 26;8(4):706. doi: 10.3390/vaccines8040706. Vaccines (Basel). 2020. PMID: 33255895 Free PMC article. Review.
-
Plasmacytoid Dendritic Cells and Cancer Immunotherapy.Cells. 2022 Jan 11;11(2):222. doi: 10.3390/cells11020222. Cells. 2022. PMID: 35053338 Free PMC article. Review.
-
Immune adjuvant efficacy of CpG oligonucleotide in cancer treatment is founded specifically upon TLR9 function in plasmacytoid dendritic cells.Cancer Res. 2011 Oct 15;71(20):6428-37. doi: 10.1158/0008-5472.CAN-11-2154. Epub 2011 Jul 25. Cancer Res. 2011. PMID: 21788345 Free PMC article.
-
Distinctive CD8+ T cell activation by antigen-presenting plasmacytoid dendritic cells compared to conventional dendritic cells.Cell Rep. 2025 Mar 25;44(3):115413. doi: 10.1016/j.celrep.2025.115413. Epub 2025 Mar 11. Cell Rep. 2025. PMID: 40073016
-
Organ-dependent in vivo priming of naive CD4+, but not CD8+, T cells by plasmacytoid dendritic cells.J Exp Med. 2007 Aug 6;204(8):1923-33. doi: 10.1084/jem.20062373. Epub 2007 Jul 23. J Exp Med. 2007. PMID: 17646404 Free PMC article.
Cited by
-
Developing dendritic cell for SARS-CoV-2 vaccine: Breakthrough in the pandemic.Front Immunol. 2022 Sep 6;13:989685. doi: 10.3389/fimmu.2022.989685. eCollection 2022. Front Immunol. 2022. PMID: 36148241 Free PMC article. Review.
-
The role of dendritic cells in cancer and anti-tumor immunity.Semin Immunol. 2021 Feb;52:101481. doi: 10.1016/j.smim.2021.101481. Epub 2021 May 20. Semin Immunol. 2021. PMID: 34023170 Free PMC article. Review.
-
The Potential of Exosomes in Allergy Immunotherapy.Vaccines (Basel). 2022 Jan 17;10(1):133. doi: 10.3390/vaccines10010133. Vaccines (Basel). 2022. PMID: 35062793 Free PMC article. Review.
-
DC-Based Vaccines for Cancer Immunotherapy.Vaccines (Basel). 2020 Nov 26;8(4):706. doi: 10.3390/vaccines8040706. Vaccines (Basel). 2020. PMID: 33255895 Free PMC article. Review.
-
The roles of extracellular vesicles in the immune system.Nat Rev Immunol. 2023 Apr;23(4):236-250. doi: 10.1038/s41577-022-00763-8. Epub 2022 Aug 4. Nat Rev Immunol. 2023. PMID: 35927511 Free PMC article. Review.
References
-
- Steinman R. M., Some interfaces of dendritic cell biology. APMIS 111, 675–697 (2003). - PubMed
-
- Colonna M., Trinchieri G., Liu Y. J., Plasmacytoid dendritic cells in immunity. Nat. Immunol. 5, 1219–1226 (2004). - PubMed
-
- Gilliet M., Cao W., Liu Y. J., Plasmacytoid dendritic cells: Sensing nucleic acids in viral infection and autoimmune diseases. Nat. Rev. Immunol. 8, 594–606 (2008). - PubMed
-
- Alculumbre S. G. et al. ., Diversification of human plasmacytoid predendritic cells in response to a single stimulus. Nat. Immunol. 19, 63–75 (2018). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
